Dras. MŞ Jose Broseta Prades y Elvira Smith Ferres.
Departamento de Anatomía. Facultad de Medicina y
Odontología (UNIVERSIDAD DE VALENCIA).
AMIGDALA: SINDROME DE KLÜVER-BUCY
Juan Emilio López Centelles.
Valencia a 23 de abril del 2008
Índice
- 1.- INTRODUCCIÓN AL SISTEMA LÍMBICO EMOCIONAL
- 1.1.- Perspectiva histórica
- 1.2.- Componentes
- 1.3.- Función
- 2.- AMÍGDALA: SÍNDROME DE KLÜVER – BUCY
- 2.1.- Amígdala (núcleos, aferencias y eferencias)
2.2.- Síndrome de Klüver-Bucy
- - Definición
- - Causas
- - Cuadro clínico
- - Pronóstico y tratamiento
- BIBLIOGRAFÍA
Agradecimientos
Debo agradecer este trabajo a mi madre, por ayudarme a buscar la
biografía, y también a mis profesoras de neuroanatomía, por facilitarme
la tarea a la hora de realizar este trabajo.
Palabras Clave
Límbico, Amígdala, Síndrome, Klüver-Bucy, Demencia.
1.- INTRODUCCIÓN AL SISTEMA LÍMBICO EMOCIONAL
1.1.- PERSPECTIVA HISTÓRICA
Los primeros estudios sobre el sistema límbico fueron desarrollados en
el siglo XIX por el neurólogo francés Pierre Paul Broca, quien fue el
primero seńalar las relaciones existentes entre las emociones y lo que
denominó “le grande lobe limbique”, haciendo referencia a las
estructuras que rodeaban al diencéfalo y bordeaban así el córtex
(formación hipocampal, amígdala, circunvoluciones del cíngulo y
parahipocampal).En el mismo siglo, el neuroanatomista alemán Alois
Alzheimer observó la aparición de ovillos neurofibrilares y placas de
betaminoide en estas regiones asociadas a la demencia, estos cambios
eran especialmente notables en la formación hipocampal. Una vez
asentados estos conocimientos se establecieron relaciones entre el
lóbulo límbico y funciones cognitivas como el pensamiento, la memoria y
aspectos de nuestra personalidad. En 1937, James Papez, neuroanatomista
de la universidad de Cornell, descubrió el circuito hipocampo-mamilo-
talámico, y lo relacionó con los sistemas emocionales del cerebro, y
más adelante se vio que muchas de las estructuras identificadas por
Papez jugaban un papel importante en el aprendizaje, la memoria y la
cognición. (Martin, 2001, p.447).
1.2.- COMPONENTES
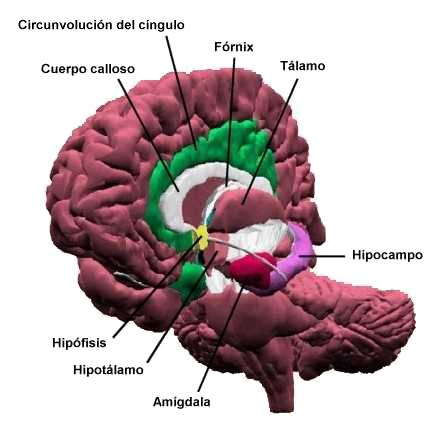
El concepto de sistema límbico se puede desarrollar desde dos puntos de
vista diferentes. Desde el punto de vista estructural está compuesto
por el lóbulo límbico, comprendiendo por el área subcallosa
(circunvoluciones paraolfatoria y paraterminal), la circunvolución
cingular, el istmo de la circunvolución cingular, la circunvolución
parahipocámpica y el uncus, el lóbulo límbico también incluye la
formación hipocámpica que queda en profundidad (ver fig. 1A). Desde el
punto de vista funcional comprende, además de las estructuras del
lóbulo límbico, núcleos y fascículos subcorticales, que en su conjunto
forman el sistema límbico. Estos núcleos son los núcleos septales y el
núcleo accumbens, varios núcleos del hipotálamo especialmente aquello
asociados al tubérculo mamilar, los núcleos del complejo amigdalino y
la sustancia innominada adyacente, y partes de tálamo, particularmente
los núcleos anteriores y dorsomedial. Otras estructuras conectadas con
el sistema límbico son los núcleos de la habénula, el área tegmentaria
ventral y la sustancia gris periacueductal. Otros estudiosos consideran
a la corteza prefrontal como un componente importante el sistema
límbico debido a su capacidad de influir sobre otras regiones
corticales y subcorticales de dicho sistema (Haines, 2007, p.494). Los
fascículos subcorticales comprenden haces de fibras eferentes del
sistema límbico como el fornix, la estría terminal, y la vía
amigdalofuga ventral y el fascículo mamilofalámico de Vicq d’Azyr. (Ver
fig. 1B y 2).
1.3.- FUNCIÓN
Pese a que los estudiosos de esta materia no han llegado a un consenso
sobre las funciones del sistema límbico se sabe que influye en los
sistema neuroendocrino, regulación autónoma y motor somático. Además
existen dos estructuras subcorticales clave, la formación hipocampal y
la amígdala y cada una de ellas forma circuitos distintos y complejos
con el resto del encéfalo. Los circuitos hipocampales están formados
por distintas estructuras diencefálicas y telencefálicas, estos
circuitos son esenciales para consolidad la memoria a corto plazo en
memoria a largo plazo y para la memoria espacial, así veremos como en
el síndrome de Korsakoff, en el que se alteran dichos circuitos,
aparecen graves disfunciones neurológicas, como la pérdida de memoria.
Los circuitos de la amígdala al parecer intervienen básicamente en las
emociones y en sus expresiones conductuales como la furia, más adelante
veremos que con la perdida bilateral de esta estructura aparecen
alteraciones conductuales que constituyen el síndrome de Klüber-Bucy.
(Martin, 2001, p. 448).
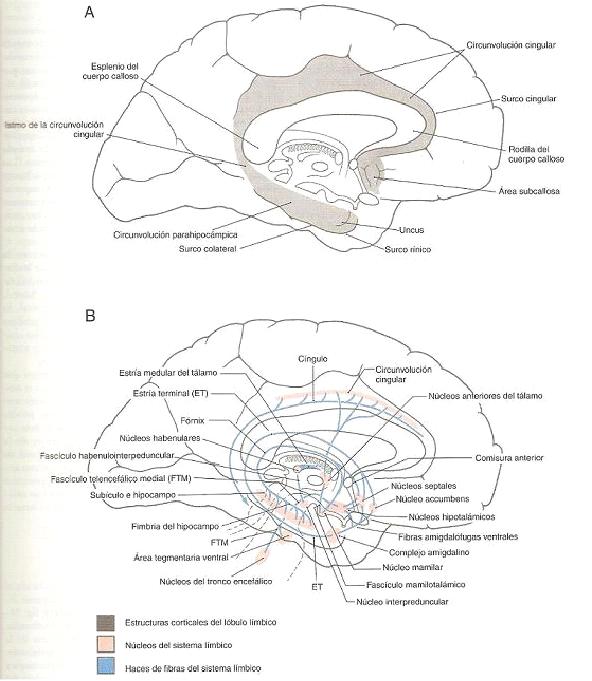
Fig. 1. Imagen anatómica de lóbulo límbico y las
circunvoluciones que lo componen (A). Esquema de las principales
estructuras que integran el sistema límbico con sus principales núcleos
y fascículos (B). (Haines, 2007, p.495).
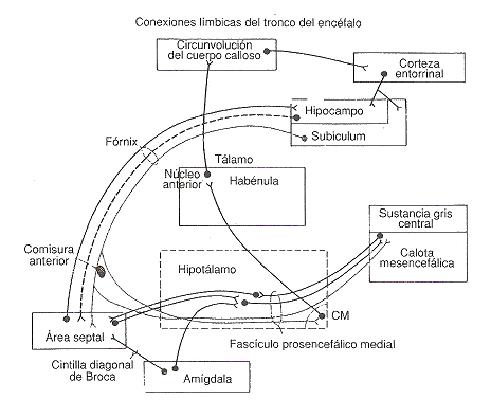
Fig. 2. Esquema de las principales conexiones entre las
diversas estructutras del sistema límbico. (Carpenter, 1994, p.359).
2.- AMÍGDALA: SÍNDROME DE KLÜVER – BUCY
2.1.- Complejo amigdalino (estructura, aferencias y eferencias).
- Introducción
La amígdala (ver fig.3 y 4) es clave para nuestras experiencias
emocionales. De esta estructura subcortical dependen los estímulos a
los que respondemos, el modo en que están organizadas las respuestas
que manifestamos a estos estímulos así como las respuestas internas de
los órganos de nuestro cuerpo. Es como un gran integrador de aferencias
sensitivas para modular respuestas efectoras en relación al medio
externo e interno. En primates no humanos, tras lesión de la amígdala,
los objetos que representaban una amenaza dejan de provocar miedo, o no
se distinguen los objetos no comestibles de los comestibles. Por el
contrario, la estimulación eléctrica de la amígdala intacta según el
lugar concreto en el que se aplique evoca en los carnívoros diversas
reacciones viscerales y de defensa. (Martin, 2001, p.457).
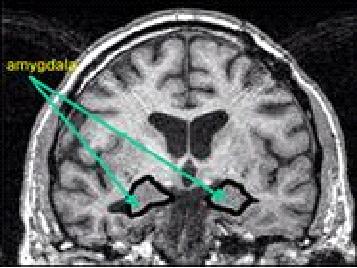
Fig. 3. Localización anatómica de la amígdala vista en corte
coronal en una resonancia magnética. (Möller T.B., 1996, p. 125)
- Estructura
El complejo amigdalino es un grupo de células nerviosas que se
encuentran en la parte dorsomedial del lóbulo temporal, profundo al
uncus. Queda inmediatamente rostral a la formación hipocámpica y al
extremo anterior del asta temporal del ventrículo lateral. El complejo
amigdalino está compuesto por varios núcleos que se sistematizan para
su estudio en un grupo basolateral y un grupo corticomedial (incluido
el núcleo central). El grupo basolateral tiene interconexiones con
estructuras corticales y el grupo corticomedial se relaciona más
íntimamente con el olfato. (Haines, 2007, p. 500).
De las aferencias y eferencias que a continuación nombramos se puede
extraer las diversas funciones que realizarán los diferentes núcleos.
Los núcleos basolatelales, asignan importancia emocional a los
estímulos, sus conexiones al hipocampo parecen ser importantes para
aprender el significado emocional de estímulos complejos o de su
contexto (martin, 2001, p. 457. ). El núcleo central recibe información
que llega desde el interior del cuerpo para modular la respuesta
visceral. Finalmente los núcleos corticomediales reciben información
olfatoria y regulan conductas relacionadas con el olfato como la sexual
o el apetito dependiendo de los olores percibidos (Martin, 2001, p.
458.).
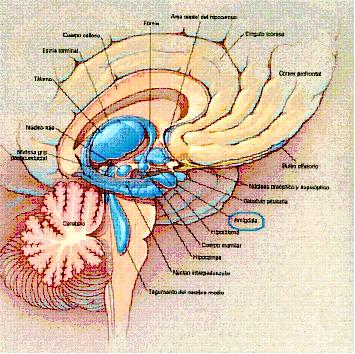
Fig. 4. Localización tridimensional de la formación amigdalina
en el cerebro humano. (Nolte J., Ph.D. 1995, p. 253)
- Aferencias (ver fig. 5A)
El grupo basolateral recibe aferencias del tálamo, la corteza
prefrontal, las circunvoluciones cingular y parahipocámpica del lóbulo
temporal, la corteza de la ínsula y el subículo. Estas fibras
proporcionan información somatosensorial, visual y visceral al complejo
amigdalino. El grupo corticomedial recibe información olfativa, fibras
desde el hipotálamo (núcleo ventromedial, área hipotalámica lateral) y
desde los núcleos dorsomedial y medial del tálamo. Además este grupo
celular, en particular este núcleo central, recibe información
ascendente desde los núcleos del tronco del encéfalo que participan en
las funciones viscerales, como los núcleos parabraquiales, el núcleo
solitario y parte de la sustancia gris periacueductal, entre otros.
(Haines, 2007, p.501).
- Eferencias (ver fig. 5B)
Las dos vías principales del complejo amigdalino son la estría terminal
y la vía amigdalofuga ventral. La estría terminal es una haz de fibras
que se origina en las células del grupo corticomedial, durante la mayor
parte de su trayecto, este haz se encuentra en el surco entre el
núcleo caudado y el tálamo, acompańado por la ven terminal, a lo largo
de su longitud se asocia a unos agregados discontinuos de células que
en conjunto se denominan núcleo del lecho de la estría terminal. Este
haz se distribuye por varios núcleos del hipotálamo (los núcleos
preópticos, el ventromedial, el anterior y al área hipotalámica
lateral), el núcleo accumbens, los núcleos septales y las áreas
rostrales del núcleo caudado y del putamen. La vía amigdalofuga ventral
es el principal haz de fibras eferentes del complejo amigdalino. Esto
axones se originan tanto en el grupo basolateral como en el núcleo
central del grupo celular corticomedial, y siguen dos trayectorias
generales. Los axones que provienen en su mayor parte de las células
basolaterales adoptan una disposición medial a través de las sustancia
innominada (en la cual terminan algunas de sus fibras) para finalmente
hacer sinapsis en el hipotálamo y los núcleos septales. La sustancia
innominada da origen a una proyección colinérgica difusa hacia la
corteza cerebral. Es probable que estas fibras tengan un cometido en la
activación de la corteza cerebral como respuesta a estímulos
importantes desde el punto de vista conductual. Asimismo, las células
del grupo basolateral también proyectan de forma difusa hacia las
cortezas prefrontal, cingular, insular y temporal inferior. Otras
fibras, sobre todo las del núcleo central, giran en sentido caudal y
descienden de manera difusa por el tronco del encéfalo para terminar en
los núcleos viscerales (motor dorsal del vago), núcleos del rafe
(magno, oscuro, pálido) y otras áreas como el locus ceruleus, los
núcleos parabraquiales, y la sustancia gris periacueductal. La mayoría
de estas áreas del tronco del encéfalo que reciben información desde la
amígdala proyectan de nuevo hacia esta estructura. Otra ruta a través
de la cual las proyecciones del hipocampo y la amígdala influyen sobre
el tronco del encéfalo es la estría medular del tálamo. Este haz
transporta fibras de los núcleos septales (destino de las conexiones
amigdalinas e hipocámpicas) hacia los núcleos habenulares. A su vez
este último grupo de células da origen al fascículo
habenulointerpeduncular, que proyecta hacia al núcleo interpeduncular y
a otros puntos del mesencéfalo, como el área tegmentaria ventral y la
sustancia gris periacueductal. (Haines, 2007, p. 501)
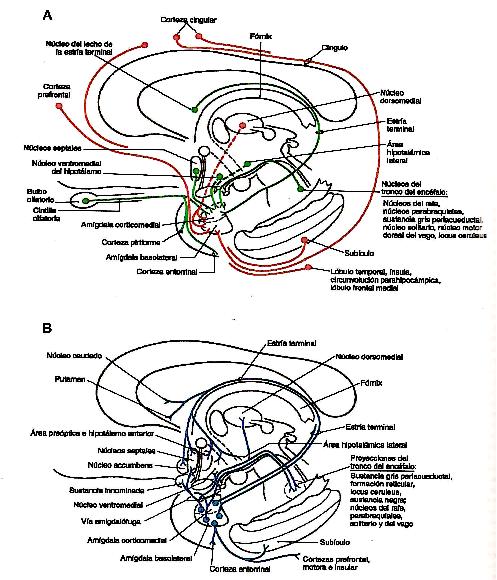
Fig. 5. Representación esquemática de las aferencias y
eferencias del complejo anigdalino. (Haines, 2007,
p.502).
3.2.- SÍNDROME DE KLÜVER-BUCY
- Definición
El síndrome toma el nombre de Heinrich Klüver y Paul Bucy que en 1.937
extrajeron los lóbulos temporales a varios monos para determinar la
función de esta parte del cerebro. Las lesiones bilaterales del lóbulo
temporal que destruyen gran parte del complejo amigdalino producen una
serie de cambios conductuales llamados síndrome de Klüver-Bucy. Este
déficit se describió inicialmente en una serie de experimentos con
animales, pero también se ha observado en pacientes como consecuencia
de un traumatismo en el lóbulo temporal o de una intervención
quirúrgica contra la epilepsia en esa zona. El dańo del complejo
amigdalino frecuentemente afecta porciones de las estructuras vecinas y
de la sustancia blanca que lo rodea, y su extensión hacia otras
estructuras puede contribuir al cuadro clínico. La lesión de la
amígdala y el hipocampo produce un déficit de memoria mayor que el que
se produce con el dańo de cada una de estas estructuras por separado.
- Causas
La incidencia en humanos del síndrome suele ocurrir típicamente como
resultado de:
- - Lesiones quirúrgicas
- - Meningoencefalitis
- - Enfermedad de Pick: Demencia presenil progresiva caracterizada
por atrofia cerebral con predominio en los lóbulos frontal y temporal.
(Masson, 2001, p.402).
- - Encefalopatía anóxico-isquémica
- - Encefalitis por herpes simple
- - Neurocisticercosis
- - Traumatismo craneoencefálico con gliosis
- - Meningitis tuberculosa.
- Cuadro clínico
En general se puede decir que los pacientes con este transtorno son
mansos, pueden comer objetos no apropiados, presentan hiperoralidad
irrefrenable, y a veces contrastan con hipersexualidad, pérdida del
miedo, con irritabilidad e ira, todo esto vivido con placer.
Los síntomas típicos incluyen:
- Agnosia visual y puede también exhibir agnosia táctil y auditiva.
Este signo puede desdoblarse en astereognosia y prosopagnosia.
- Hiperoralidad o a olerlos, esto incluye objetos peligrosos o dańinos,
como una cerilla encendida.
- Hipermetamorfosis y reaccionar exageradamente a los estímulos
visuales.
- Placidez o apacibilidad.
- Hiperfagia aunque no tenga hambre, o comer objetos que no son
alimentos o alimentos inadecuados para su especie.
- Hipersexualidad incluyendo conversaciones impúdicas, vagos intentos
de contacto sexual y actividad autoerótica enormemente aumentada.
- Además de estos problemas previsibles los pacientes también pueden
experimentar amnesia, demencia o afasia, dependiendo de la extensión de
la lesión en el lóbulo temporal.
- Pronóstico y tratamiento
El pronóstico del síndrome de Klüver-Bucy es malo y la recuperación es
muy escasa en la mayoría de los pacientes, especialmente cuando la
patología originaria es la encefalitis herpética, que deja lesiones
irreversibles, o procesos metabolico-degenerativos progresivos. Hay
descripciones de presentación aguda del síndrome de Klüver-Bucy, la
mayoría de las veces tras un traumatismo craneal que afecta a ambos
lóbulos temporales, cuya sintomatología mejora considerablemente o
incluso llega a desaparecer tras un tiempo prudencial, que puede ir
desde unos días hasta unos meses e incluso 1 ańo, habiéndose
relacionado, en algunos casos, la mejoría con tratamiento farmacológico
con carbamazepina. Sin embargo, otros pacientes con síndrome de Klüver-
Bucy postraumático también se han recuperado de las manifestaciones del
síndrome en un tiempo muy breve con la administración de otras
sustancias, tales como fenobarbital y benzodiazepinas.
Bibliografía
BIBLIOGRAFÍA ESENCIAL
Cambier J., Masson M., Dehen H. Manual neurología. Barcelona:
Masson.
Carpenter. 1994. Neuroanatomia fundamentos. Madrid: Editorial
médica
panamericana S.A.
Haines D.E.. 2007. Principios de neurociencia. Madrid: Elsevier.
Harrison. 1998. Principios de medicina interna. Madrid: McGraw-
Hill.
Klüver H, Bucy PC: Preliminary analysis of functions of the temporal
lobes in monkeys. Arch Neurol Psychiatry 1939; 42:979–1000
Martin J.H. 2001. Neuroanatomía. Madrid: Prentice Hall.
Möller T.B., Reif E., 1996. Atlas de bolsillo de cortes anatómicos:
TC y RM. Madrid: Editorial Médica Panamericana
Nolte J., Ph.D. 1995. El cerebro humano: introducción a la anatomía
funcional. Madrid: Mosby / Doyma Libros.
Stewart JT: Carbamazepine treatment of a patient with Klüver-Bucy
syndrome. J Clin Psychiatry 1985; 46:496–497
Glosario de términos
- Agnosia visual: pérdida de la capacidad para reconocer objetos con la
vista.
- Astereognosia: incapacidad para reconocer objetos.
- Prosopagnosia: incapacidad para reconocer caras.
- Hiperoralidad: tendencia en exceso a examinar los objetos con la
boca.
- Hipermetamorfosis: compulsión de explorar a fondo el ambiente
cercano.
- Hiperfagia: el paciente puede comer excesivamente.
- Hipersexualidad: aumento de la conducta sexual.
Apéndice Biografico
- Paul Bucy, neuroanatomista, nació 13 de noviembre de 1904 en Hubbard,
Iowa y falleció el 22 de septiembre de 1992 en su ciudad natal.
- Heinrich Klüver, psicólogo, hijo de Wilhelm y Dorothes (Wübbers)
Klüver, nació el 25 de mayo de 1897 en Schleswig-Holstein, Alemania y
falleció el 8 de febrero de 1979 en Oak Lawn, Illinois, a la edad de 81
ańos.