
UNIVERSIDAD DE VALENCIA
Facultad de Medicina y Odontología
Diseńo y presentación de trabajos científicos
MŞ Alejandra Dubois Marques
Sistema Endocrino:
Pieza básica para la adaptación
Resumen
El sistema endocrino se basa en la síntesis de hormonas con el fin de regular el medio interno del organismo; así en este documento se presentan tres elementos importantes
que forman parte del mismo.La médula suprarrenal es un ganglio simpático agrandado y especializado que sintetiza adrenalina y noradrenalina; estas son estimuladas en respuesta
a la actividad del SN simpático, en casos de hipoglucemia, hipovolemia, hipotensión, ejercicio o estrés. La adrenalina aumenta la glucosa en plasma, los ácidos grasos libres y
los cetoácidos, en cambio la noradrenalina tiene efectos sobre acciones cardiovasculares como por ejemplo aumentar la frecuencia cardíaca; muchas veces la adrenalina es reforzada
por la noradrenalina como neurotransmisor.
La glándula tiroides se encarga de la síntesis de T3 y T4; la T4 actúa en gran medida como prohormona, dando lugar a T3, que es la principal hormona activa. Esta hormona
se encarga de aumentar la tasa metabólica basal entre otras acciones de destacada importancia.
Los islotes pancreáticos secretan principalmente insulina, glucagón y somatostatina; estas tres hormonas tienen efectos diferentes, así la insulina se encarga de disminuir los
niveles plasmáticos de glucosa y almacenarla en forma de glucógeno; el glucagón se encarga de aumentar los niveles plasmáticos de glucosa, ácidos grasos y cetoácidos; y la
somatostatina coordina la llegada de nutrientes con la utilización de sustratos, entre otras.
Palabras clave
Catecolaminas, adrenalina, noradrenalina, T3, T4, TSH, TRH, retroalimentación, insulina, glucagón, somatostatina, hiperglucemia.
Indice
- Médula suprarrenal
- Síntesis, almacenamiento y metabolismo
- Regulación
- Acciones de las hormonas catecolaminas
- Mecanismos intracelulares
- Efectos sobre el organismo
- Integración de la respuesta al estrés
- Tiroides
- Síntesis y secreción
- Regulación
- Metabolismo
- Acciones de las hormonas tiroideas
- Mecanismo intracelulares
- Acciones sobre el organismo
- Páncreas endocrino
- Insulina
- Síntesis y secreción
- Regulación
- Acciones de la insulina
- Glucagón
- Síntesis y secreción
- Acciones del glucagón
- Índice insulina/glucagón
- Somatostatina
- Bibliografía
Introducción
El sistema endocrino es una pieza básica de la adaptación del organismo humano a las alteraciones del ambiente interno y externo. Este sistema mantiene el medio interno estable
a pesar de las variaciones en las entradas o salidas de sustratos, minerales, agua, moléculas ambientales, calor y otros factores. De esta forma el sistema endocrino y nervioso
actúan conjuntamente para mantener la homeostasis.
Las hormonas son moléculas transmisoras de seńales transportadas por la sangre, por axones y por sangre o por difusión local. Así mismo las encontramos de diferente naturaleza
pudiendo ser proteicas, peptídicas, catecolamínicas, esteroideas o derivados de yodados de la tirosina.A través de estas hormonas y de los diversosórganos del cuerpo el sistema
endocrino regulará el metabolismo, los líquidos corporales, el crecimiento y el desarrollo sexual. A continuación nos centraremos en el estudio de tres elementosfundamentales
y sus acciones sobre el organismo, estos son: la médula suprarrenal, la glándula tiroides y el páncreas endocrino.
Desarrollo
La médula suprarrenal ocupa la porción interna de la glándula suprarrenal y se encarga de la síntesis de catecolaminas principalmente de adrenalina y noradrenalina.Suele
activarse a la vez que la Proción simpática del SN autónomo y actúa de acuerdo con él en la reacción de “lucha o huida”
Síntesis, almacenamiento y metabolismo
Las catecolaminas se metabolizan en el hígado y se secretan por el rińón. Derivan de la tirosinaque se convierte en DOPA que pasa a dopamina y posteriormente a noradrenalina.
Parte de esta es almacenada en gránulos y otra se difunde al citoplasma donde es convertida en adrenalina que se almacena de nuevo en gránulos. Este almacenamiento se produce
gracias a la presencia de ATP; incluso muchas veces se utiliza analizar los niveles de ATP en sangre para saber el de catecolaminas (Robert; Matthew, 1999) ya que es más fácil.
La estimulación simpática aguda de la médula activa el paso de tirosina a DOPA*, al igual que la estimulación crónica que mantiene la producción de
catecolaminas. El cortisol induce el paso de noradrenalina a adrenalina con la ayuda del SN simpático (Robert; Matthew, 1999), de modo que estimula su síntesis.
Toda la adrenalina circulante es segregada por la médula, en cambio toda la noradrenalina circulante procede de las terminaciones nerviosas simpáticas y del encéfalo (hendiduras
sinápticas) (Linda, 2000). Tienen una vida media de 2min y ambas son metabolizadas en el hígado y el rińón por dos enzimas que permiten obtener como productos finales AVM* y
metanefrinas que se secretan por la orina (Robert; Matthew, 1999). Sirven como indicadores de los niveles de catecolaminas en sangre, y de la actividad del SNS o de una hipersecreción
patológica de adrenalina.
Inicio
Regulación
La secreción de la médula forma parte de la reacción de lucha o huida, por tanto situaciones de estrés producen una rápida secreción de adrenalina y noradrenalina. Estos estímulos
son captados por el SN Simpático e inician respuestas en el hipotálamo y el encéfalo. Sin embargo, la secreción de adrenalina aumenta específicamente en respuesta a hipoglucemias
leves, hipoxia* moderada y ayuno aunque la actividad del SN sea constante (Robert; Matthew, 1999). La hipoglucemia apenas modifica las concentraciones de noradrenalina; la disminución
de la presión venosa aumenta los niveles de ambas, pero sólo la concentración de adrenalina es lo bastante elevada para aumentar la frecuencia cardíaca y la presión arterial. La
adrenalina funciona así como hormona, mientras que la noradrenalina no; esta última contribuye como neurotransmisor a las respuestas compensadoras frente a la hipovolemia* y a
hipoglucemias* severas, ya que la mayor concentración necesaria para su acción se alcanza en la zona efectora.
Inicio
Acciones de las hormonas catecolamínicas
Mecanismos intracelulares
Estas catecolaminas ejercen muchos de sus efectos a través de receptores de la membrana plasmática, designados como a1, a2, ß1, y ß2. Los receptores ß1, ß2 están
acoplados a la proteína G estimulante de la adenilciclasa, aumentando los niveles de AMPc; en cambio el a2 está acoplado a la proteína G inhibidora de la adenilciclasa. De
este modo las catecolaminas pueden poner en marcha (ß1, ß2) o desactivar (a2) una cascada de fosforilación de proteínas. El a1 emplea como segundos mensajeros el Ca+2 y
los derivados del IP3 (Linda, 2000). Además según donde actúe, el receptor es constrictor o dilatador y en muchos casos depende del predominio de los mismos (la
constricción en piel se debe básicamente a receptores a1). La exposición crónica a catecolaminas disminuye el número de receptores. Así las catecolaminas presentan un tipo de
retroalimentación negativa rápida (Wikipedia, 2008) que limita casi inmediatamente las acciones hormonales
Efectos sobre el organismo
- Aumento de la glucogenólisis: Estimulada por la activación mediada por AMPc
- Aumento de la resistencia periférica, aumento de la presión arterial; ya que provocan contracción
- Aumento del gasto cardíaco
- Aumento del metabolismo basal
- Disminución de la movilidad y secreciones intestinales
- Broncodilatadores
- Estimulantes del SNC
Una técnica de relajación utilizada es aumentar la secreción salival ya que de este modo disminuye la frecuencia cardíaca (es un estímulo del SN parasimpático)
Integración de la respuesta al estrés
La médula y la corteza suprarrenal son dos elementos importantes en la adaptación al estrés; así se establece una íntima relación entre el SN simpático y el eje
hormona liberadora de CRH y ACTH (Robert; Matthew, 1999). Las situaciones de estrés intenso como anticipación de peligros, miedo, excitación, traumatismos,
hipovolemia, hipotensión, hipotermia, hiperglucemia, ejercicio intenso…, activan casi simultáneamente las hormonas productoras de CRH y las neuronas adrenérgicas del
hipotálamo. Esta activación se refuerza mutuamente, ya que la noradrenalina aumenta la liberación de CRH y la CRH eleva los niveles plasmáticos de cortisol; la estimulación
adrenérgica eleva los niveles de catecolaminas. Juntas ambas hormonas aumentan la producción de glucosa y desplazan su utilización al SNC (Robert; Matthew, 1999), así
mismo también aumentan la presión arterial y el gasto cardíaco y mejoran el aporte de sustratos a los tejidos para una defensa inmediata.
La estimulación adrenérgica produce una activación de la conducta útil para la defensa y una oportuna agresividad. Al mismo tiempo, la CRH actúa sobre otras
hormonas hipotalámicas inhibiendo la liberación de la GH y de gonadotropinas, ya que el crecimiento y la reproducción no son funciones útiles durante el estrés (Linda,
2000). El cortisol por su parte inhibe el crecimiento y la ovulación (regula la producción de glucocorticoides a través de la hipófisis) y la CRH la actividad sexual y la
ingesta. Así se demuestra la integración del sistema nervioso y endocrino.
El estrés excesivo puede tener efectos perjudiciales como hipertensión arterial, obesidad, infecciones, diabetes (Wikipedia, 2008). Existen dos tipos de estrés:
Estrés del luchador:
- Predomina un estímulo simpático.
- Secreción de adrenalina, noradrenalina, ß-endorfina (efectos como el placer), prolactina, renina y oxitocina
- Va seguido de diferentes actividades físicas (respuesta activa)
Estrés pasivo:
- Predominio corticosuprarrenal
- Tiene efectos más graves porque viene producido por el cortisol y la ACTH y además no produce ß-endorfina
Inicio
La glándula tiroides produce dos hormonas, tiroxina y triyodotironina, a un ritmo más o menos constante. Estas aumentan la tasa basal de utilización de O2 y la
tasa metabólica basal, así como la tasa de producción de calor (Robert; Matthew, 1999). Sus acciones son básicas para el crecimiento y la maduración del feto y del nińo. Se
sitúa delante de la tráquea. Su estructura presenta células que están rodeadas por una membrana basal y forman folículos (unidad funcional) circulares de capa única. La luz
de los folículos contiene hormonas tiroideas almacenadas en un material coloide.
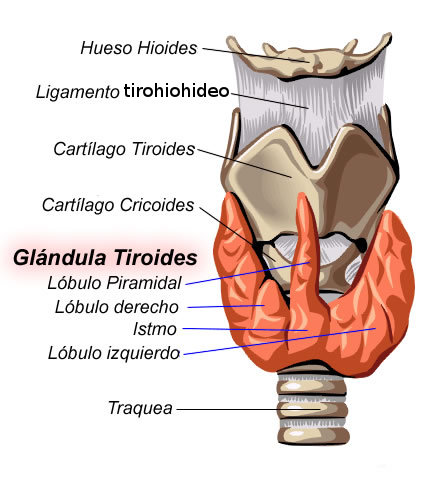
Fig. Nş 1. Glándula tiroides y sus límites (Wikipedia, 2008)
Síntesis y secreción
Son las únicas hormonas que incorporan un elemento inorgánico, el yodo, en una estructura orgánica formada por dos moléculas de tirosina. El principal producto es la tiroxina (T4) que
actúa como prohormona circulante, y en cantidad menor la triyodotironina (T3) que ejerce actividad hormonal en las células diana (Schwartz, 1983) y se produce a partir de la T4.
Estas moléculas a su vez van unidas a una proteína denominada tiroglobulina (sustancia realmente yodada), de aquí que su liberación requiera hidrólisis proteolítica.
El yoduro es un elemento dietético esencial debido a su función tiroidea. Pequeńos aumentos de la ingesta dietética del mismo aumentan la tasa de síntesis de hormonas
tiroideas. Sin embargo, si la ingesta es muy alta se inhibe el atrapamiento de más yoduro y el mecanismo biosintético, disminuyendo la producción hormonal hasta la que se normalice.
Una vez yodada la tiroglobulina, se almacena en el folículo en forma de coloide. La liberación a la sangre de T3 y T4 requiere que la tiroglobulina pase al interior de la
célula endocrina por endocitosis (Wikipedia, 2008); la membrana de la célula forma seudópodos que engloban una porción del coloide que posteriormente se separa
formando una gotita que se desplaza en dirección basal. Ahí las proteasas lisosomales liberan T3 y T4 que pasan a la sangre capilar adyacente.
Inicio
Regulación
El principal estímulo de la secreción de hormona tiroidea es la tirotropina u hormona estimuladora del tiroides (TSH), segregada por la hipófisis. El estímulo
directo de la secreción de TSH es la hormona liberadora de tirotropina (TRH) del hipotálamo (Scwartz, 1983). Las hormonas tiroideas inhiben por retroalimentación
negativa la síntesis y liberación de TSH y TRH.La estimulación prolongada de TRH aumenta también la síntesis de TSH y su bioactividad. La TRH disminuye con el tiempo
el número de sus propios receptores con lo que la hormona liberadora pierde su eficacia.La TSH está formada por dos subunidades peptídicas. La subunidad a es inespecífica,
ya que forma parte también de tres hormonas diferentes relacionadas con la reproducción; en cambio la subunidad ß debe combinarse con la subunidad a para que la TSH
estimule las células tiroideas. Esta hormona actúa sobre las células foliculares de la glándula, produciendo numerosos efectos como: atrapamiento de yoduro y todos los
pasos de la síntesis de T3 y T4, así como la endocitosis de coloide y la liberación proteolítica de T3 y T4 por la glándula (Linda, 2000). La exposición crónica a la TSH
provoca hiperplasia de las células foliculares. En ausencia de TSH, la glándula se atrofia aunque sigue manteniendo un nivel basal de secreción.
Control por retroalimentación
La producción de hormonas se halla bajo un fino control por retroalimentación, que mantiene los niveles de T3 y T4 constantes. Cambios en los niveles de hormona tiroidea de un 10-30%
bastan para desplazar los niveles de TSH en sentido contrario. El retrocontrol negativo se ejerce principalmente a nivel hipofisario.La molécula responsable es la T3 generada por
desyodación, la cual suprime la síntesis y liberación de TSH, disminuye también los receptores de TRH para reducir la TSH (Robert; Matthew, 1999).La TSH también es inhibida por
la dopamina y somatostatina, así como también el cortisol y la GH.
Inicio
Metabolismo
La T4, principal forma circulante y segregada, actúa como prohormona de la T3 y tiene una vida media prolongada de 6 días. La mayor parte de T3 procede de la
desyodación de la T4 circulante, el metabolito activo se encuentra en una cantidad mucho menor y tiene una vida media más corta, de 1 día.
Unión a proteínas
Ambas hormonas circulan ligadas a una proteína denominada globulina de unión a tiroxina (TBG). Cada molécula de TBG se une a una de T4; el 70% está ligada a
esta, el resto están ligadas a la transtiretina que tiene menor afinidad (Linda, 2000). Ambas tienen dos funciones:
- Crear un reservorio circulante de T4 amortiguando los cambios del funcionamiento de la glándula tiroides.
Tras la extirpación de la glándula, la concentración de T4 tardaría casi una semana en disminuir al 50%.
- La unión a proteínas impide la filtración glomerular y se excreción urinaria.
El exceso o disminución transitoria de T4 puede ser compensado con la asociación o disociación de la misma a la TBG respectivamente (Robert; Matthew,
1999). Los cambios mantenidos conducen a la disminución o aumento mantenido de las fracciones ligadas o libres (enfermedades tiroideas). Un cambio en la concentración
de TBG altera la proporción entre T4 libre y ligada, de modo que la glándula ha de aumentar o disminuir su secreción hasta llegar de nuevo al estado de equilibrio.
Vías metabólicas
El hígado, el rińón y el músculo esquelético son los principales lugares donde se degradas estas hormonas. La tasa de eliminación
de T4 es proporcional a la concentración plasmática de T4 libre.
Acciones de las hormonas tiroideas
Mecanismo intracelular
La T3 y T4 se introducen a través de transportadores dependientes de energía, donde la mayor parte de T4 es desyodada a T3. Ambas pasan al núcleo, donde la T3 se
une a un receptor nuclear con mucha mayor afinidad que la T4. El complejo receptor-T3 interaccionará con el ADN estimulando o inhibiendo la transcripción.
Acciones sobre el organismo
Tasa de metabolismo basal: el efecto más evidente es el aumento del consumo de O2 (excepto en el cerebro, gónadas y bazo), lo que conlleva un aumento de la TMB y
temperatura corporal (Robert; Matthew, 1999). Estas hormonas aumentan la actividad de la Na+-K+-ATPasa q aumenta el consumo de O2 y la producción de calor, aumenta
el número, tamańo, superficie de las membranas y ciertas enzimas respiratorias clave de las mitocondrias.
- Metabolismo: el aumento del consumo de O2 depende, en último término, de la mayor disponibilidad de sustratos para el metabolismo oxidativo. Las hormonas
tiroideas aumentan la absorción de glucosa en el intestino y potencian la acción de otras hormonas sobre la gluconeogénesis, lipolisis y proteólisis (Wikipedia, 2008).
Por tanto aceleran la respuesta metabólica ante la inanición.
- Cardiovascular y respiratorio: al aumentar el consumo de O2 se incrementa la demanda de O2 en los tejidos (Robert; Matthew, 1999); estas hormonas por tanto
elevarán el gasto cardíaco (varían las presiones aumentando el volumen sistólico y reduciendo la resistencia vascular periférica) y la frecuencia ventilatoria
para mantener un presión arterial de de O2 y de CO2 normal a pesar del aumento de la producción de éste.
- SN simpático: aumento de la sensibilidad de las catecolaminas por la capacidad de la hormona para elevar el número de receptores ß-adrenérgicos, aumentando
así el segundo mensajero de las catecolaminas, el AMPc (Schwartz, 1983).
- Crecimiento: aumenta la actividad de los condrocitos* de la placa del crecimiento óseo.Aumenta la secreción de GH (Schwartz, 1983). Su concentración excesiva
produce la resorción ósea y la aceleración del descamamiento de la piel (estimula procesos degradativos)
- SNC: Si hay deficiencia se alteran el crecimiento de la corteza cerebral y del cerebelo, la proliferación de axones*, ramificación de las dendritas* y la
mielinización*. En nińos y adultos aumenta la velocidad y amplitud de los reflejos, la vigilia, la atención, la respuesta a estímulos, la sensación de hambre,
la memoria y la capacidad de aprendizaje (Robert; Matthew, 1999).
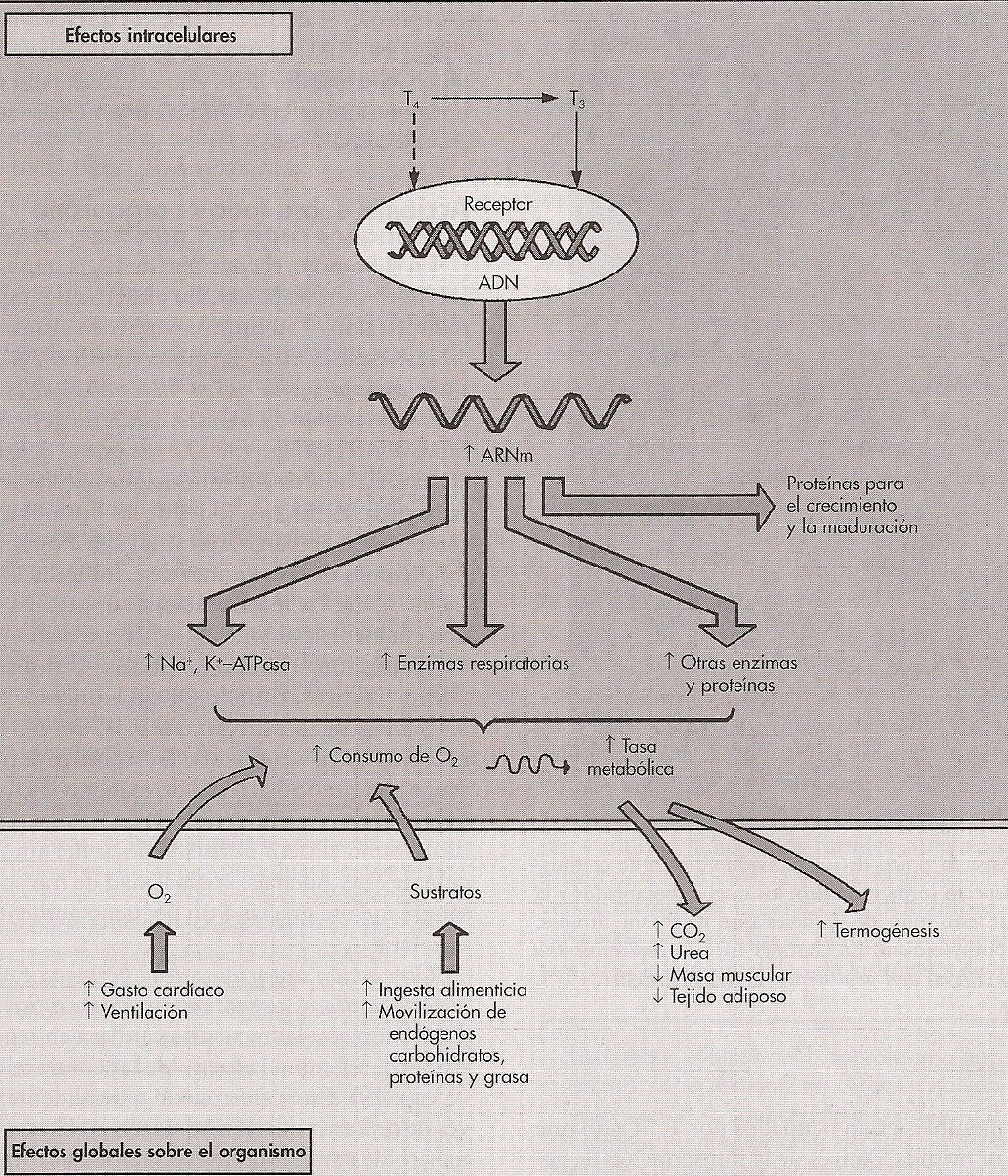
Fig. 2. Esquema general de los efectos de la hormona tiroidea. Acciones intracelulares y efectos generales sobre el organismo (Robert; Matthew, 1999)
Inicio
Las principales hormonas pancreáticas, insulina y glucagón, son rápidos y poderosos reguladores del metabolismo. Su secreción está regulada principalmente por
los niveles plasmáticos de sustratos. Juntos coordinan la disposición de los nutrientes procedentes de los alimentos, así como el flujo de sustratos endógenos mediante
acciones sobre hígado, tejido adiposo y masa muscular. Las células productoras se disponen en grupos denominados islotes de Langerhans, los cuales contienen 4 tipos de
células: las ß (65%) liberan insulina, las a (20%) liberan glucagón y las d (10%) producen somatostatina, las restantes secretan polipéptido pancreático (Robert; Matthew, 1999).
La localización de los islotes, que impone la secreción a la vena porta, hace que el hígado, órgano central del tráfico de nutrientes, esté expuesto a concentraciones de
estas hormonas más altas que los tejidos periféricos. Ambas hormonas suelen segregarse y actuar recíprocamente, cuando se necesita uno, no suele necesitarse el otro.
Síntesis y secreción
Es una hormona peptídica que contiene dos cadenas (A y B) unidas por dos puentes disulfuro. La preproinsulina contiene 4 péptidos: un péptido seńal, las cadenas A y B y un
péptido conector (péptido C) (Steiner, 1994). Cuando el péptido seńal se desdobla pasa a proinsulina la cual se dirige al RE donde se desdobla el péptido C convirtiéndose en
insulina.La insulina se segrega mediante exocitosis de sus gránulos que se disponen cerca de la membrana plasmática; cuando se aplica un estímulo los gránulos se
fusionan con la membrana liberando cantidades equivalentes de insulina y péptido C. Muchos agentes pueden estimular su secreción, pero el más importante es la glucosa.
Regulación
En sentido amplio, la secreción de insulina está gobernada por una relación de retroalimentación con el aporte de nutrientes exógenos. Si el aporte es abundante, se
segrega insulina; la hormona estimula así la utilización de los nutrientes adquiridos, al tiempo que inhibe la movilización de sustratos endógenos (Robert; Matthew, 1999).
Por el contrario si el aporte es lento o nulo, la secreción se amortigua y se estimula la utilización de nutrientes endógenos.La molécula reguladora fundamental es la
glucosa.Con niveles plasmáticos inferiores de 50mg/dl no se segrega nada o casi nada de insulina, mientras que su respuesta es máxima con niveles superiores a 250mg/dl.
Una vez completada la digestión y absorción de los nutrientes los niveles plasmáticos de glucosa y aminoácidos regresan a los niveles basales y la secreción de
insulina desciende a una tasa que se mantiene constante durante el período de ayuno nocturno habitual. Si el ayuno se prolonga durante días, la secreción de insulina declina
por debajo de la tasa basal y se reajusta a un nivel más bajo (Linda, 2000).La insulina tiene un vida media plasmática corta (6-8 horas) (Linda, 2000), debido a su degradación
específica en el rińón e hígado. Sin embargo, la misma es degradada al tiempo que ejerce sus acciones sobre las células. Con la orina se excreta muy poca insulina.
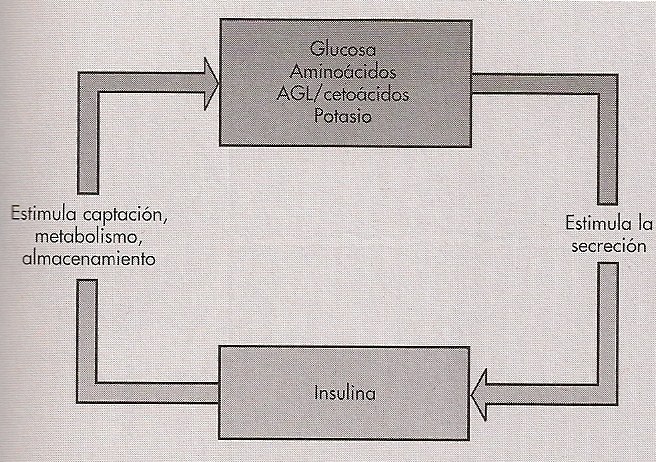
Fig. 3. Retroalimentación entre insulina y nutrientes. (Robert; Matthew, 1999)
Acciones de la insulina
- Metabolismo energético: facilita el almacenamiento de sustratos e inhibe su liberación. Los principales lugares de acción son el hígado, músculo y tejido adiposo.
- Metabolismo de los carbohidratos: en el músculo y el tejido adiposo, la insulina estimula el transporte de glucosa plasmática al citoplasma, donde es fosforilada.
En el músculo y el hígado, estimula la formación de glucógeno (Steiner, 1994). También estimula aunque en menor grado, la glucólisis y oxidación de glucosa. En el
tejido adiposo estimula la producción de a-glicerol fosfato para la formación de triglicéridos y su posterior almacenamiento.
- Metabolismo de las grasas: en el tejido adiposo, la insulina facilita la transferencia de grasas circulantes a las células adiposas al inducir la acción de una
enzima, de esta forma se liberan más ácidos grasos de los triglicéridos circulantes y son rápidamente captados por las células adiposas, donde se reesterifican
(Steiner, 1994). Así, la grasa de la dieta no necesaria para la generación inmediata de energía es almacenada. La insulina también inhibe la reacción inversa,
es decir, la lipolisis de los triglicéridos almacenados. En el hígado tiene una acción anticetogénica.
- Metabolismo de las proteínas: En el músculo estimula el transporte de ciertos aminoácidos del plasma al citoplasma (Linda, 2000). También aumenta la síntesis
global de proteínas e inhibe las enzimas proteolíticas y de la liberación de aminoácidos. La síntesis de albúmina y amilasa son estimuladas. Por tanto en medida
general contribuye al crecimiento, a la regeneración tisular y al remodelado óseo.
- Otros efectos: la translocación al espacio intracelular de proteínas, K+, PO4- y Mg+2 para la síntesis de glucógeno es estimulada por la insulina. Así como también
estimula la reabsorción de K+, PO4- y Na+ (Robert; Matthew, 1999) en los túbulos renales. Aumenta el gasto energético al estimular la bomba Na+-K+-ATPasa y
disminuye la síntesis del neuropéptido Y*.
- Mecanismos moleculares: la insulina estimula un sistema transportador de glucosa específico de la membrana de las células musculares y del tejido adiposo. Un
transportador (glut-4) (Steiner, 1994) facilita su difusión hacia el citosol. Así la insulina aumenta la síntesis de este transportador.
Correlación entre acción y secreción
Para detener la producción hepática de glucosa se necesitan concentraciones altas de insulina inducidas por la llegada de nutrientes. Las concentraciones máximas,
inducidas por una comida, estimulan mucho la captación de glucosa y aminoácidos por los tejidos periféricos, especialmente el músculo, y la captación de ácidos grasos libres
por el tejido adiposo. Este proceso permite almacenar sustratos para su futura utilización.
Inicio
Síntesis y secreción
El glucagón se sintetiza a partir de un preproglucagón en las células a de los islotes pancreáticos. Su síntesis es inhibida por niveles elevados de glucosa y estimulada por
niveles bajos de ésta. Su secreción se correlaciona, por retroalimentación, con la estimulación de la producción hepática de glucosa y el mantenimiento de los niveles
plasmáticos de la misma. Así la hipoglucemia aumenta de 2 a 4 veces la concentración de glucagón, y la hiperglucemia suprime su secreción en más del 50% (Steiner, 1994).
Estos efectos están reforzados por la insulina, la cual inhibe directamente la secreción de glucagón, y en su ausencia la estimula exageradamente. La presencia
de ácidos grasos también suprimen su liberación a diferencia de los alimentos proteicos y aminoácidos (sustratos para la producción de glucosa) que la estimulan.
El ayuno prolongado y el ejercicio mantenido aumentan la secreción de glucagón. En situaciones de estrés suele aumentar mucho también. Esto se debe probablemente
a la estimulación de las células a por el SN simpático a través de receptores a-adrenérgicos* (Robert; Matthew, 1999). Esta hormona es extraída por el
hígado en su primer paso por él y su vida media en plasma es corta. Es degradada por el hígado y el rińón y muy poca es excretada por la orina.
Acciones del glucagón
En casi todos los casos lleva a cabo acciones opuestas a las de la insulina. Favorece la movilización de glucosa, más que su almacenamiento. Tiene un efecto glucogenolítico
inmediato e intenso, impide la síntesis de glucógeno. Estimula la gluconeogénesis.Su importancia se refleja en la brusca disminución de la producción de glucosa
que se observa al inhibir su secreción (Robert; Matthew, 1999). Por el contrario, el aumento de la concentración de glucagón estimula intensamente la glucogenólisis y
aumenta con rapidez los niveles plasmáticos de glucosa. Esto ocurre incluso con niveles de insulina algo elevados.
Impide la utilización de ácidos grasos libres para la formación de triglicéridos (Steiner, 1994). Las acciones sobre el tejido adiposo y el músculo son casi insignificantes,
excepto en ausencia total de insulina. Inhibe la reabsorción tubular de Na+, lo que provoca natriuresis*.El mecanismo molecular de acción comienza con la unión a un receptor
de la membrana del hepatocito. El complejo receptor-glucagón provoca un aumento de AMPc* intracelular que va seguido de una cascada enzimática que activa la glucogenólisis (Steiner, 1994).
Inicio
Los flujos de sustratos son claramente muy sensibles a la cantidad relativa de insulina y glucagón. Cuando se exige la movilización y el aumento de utilización de sustratosendógenos,
este índice desciende; esto ocurre en el ayuno, en el ejercicio prolongado y en el período neonatal. El índice disminuye tanto por descenso de la secreción de insulina como por aumento
de la secreción de glucagón (Robert; Matthew, 1999). Cuando es conveniente almacenar los sustratos, tras una sobrecarga pura de carbohidratos o una comida mixta,este índice
aumenta, debido principalmente al aumento de la secreción de insulina. Una excepción se produce tras la ingesta de una comida proteica donde el índiceapenas se modifica;así la insulina
aumenta, captando aminoácidos y sintetizando proteínas en el músculo, pero también aumenta la secreción de glucagón, que previene la disminución de la producción de glucosa y la hipoglucemia.
Inicio
Esta hormona actúa en el hipotálamo como inhibidor de la secreción de GH* por la hipófisis. Su secreción es estimulada por la glucosa, aminoácidos, ácidos grasos
libres, glucagón y diversas hormonas gastrointestinales. La insulina inhibe su secreción. Tras una comida mixta, su concentración plasmática aumenta entre el 50 y
100% (Linda, 2000). Su efecto global es disminuir la tasa de digestión y absorción de nutrientes por el tracto gastrointestinal y su posterior utilización. Así este neuropéptido
inhibe la motilidad gástrica, duodenal y de la vesícula biliar; reduce la secreción de HCl*, jugo intestinal, enzimas pancreáticas…; inhibe la absorción de glucosa y
triglicéridos a través de la membrana de la mucosa intestinal y por último inhibe la secreción de insulina y glucagón.
Participa en un mecanismo de retroalimentación a través del cual la entrada de alimentos en el intestino estimula la liberación de la hormona para prevenir una
sobrecarga brusca de nutrientes (Linda, 2000). Las relaciones anatómicas entre las células de los islotes sugieren que las tres hormonas (somatostatina, insulina y
glucagón) influyen recíprocamente en la secreción de las otras. Esto permite mejorar la coordinación entre el tránsito, digestión y absorción.
Inicio
Inicio
Glosario
AMPc: adenosín monofosfato cíclico.
AVM: adenosín monofosfato cíclico.
Axones:prolongaciones protoplasmáticas de las neuronas, bastante largas generalmente, que transmiten el impulso nervioso.
Condrocitos: son células que se forman a partir de condroblasto y forman parte del tejido cartilaginoso. Producen y mantienen la matriz cartilaginosa.
Dendritas:prolongaciones protoplásmicas ramificadas, bastante cortas, de la célula nerviosa. Están implicadas en la recepción de los estímulos.
DOPA: (Dihidroxifenilalanina) Sustrato prácticamente inicial de la ruta metabólica de las catecolaminas.
GH: hormona del crecimiento.
HCl: ácido clorhídrico.
Hipoglucemias: concentración de glucosa en la sangre anormalmente baja.
Hipovolemia: disminución del volumen circulante de sangre debido a múltiples factores como hemorragia, deshidratación, quemaduras, entre otros.
Hipoxia: trastorno en el cual el cuerpo por completo (hipoxia generalizada), o una región del cuerpo (hipoxia de tejido), se ve privado del suministro adecuado de oxígeno.
Mielinización:formación vainas de mielina alrededor de los axones de las neuronas para acelerar la transmisión del impulso nervioso.
Natriuresis: disminución de los niveles de Na+ en sangre.
Neuropéptido Y: péptido supresor del apetito.
a-adrenérgicos: clase de receptores asociados a la proteína G, los cuales son activados por las catecolaminas.