UNIVERSIDAD DE VALENCIA
Facultad de Medicina y Odontología. Departamento de Anatomía
Y FISIOLÓGICAS
DEL SUEŃO
BASES ANATÓMICAS

Y FISIOLÓGICAS
DEL SUEŃO
BASES ANATÓMICAS
El sueńo es un estado fisiológico en el que el nivel de vigilancia está disminuido y el individuo reposa o descansa. Pero esta apariencia externa de depresión funcional es engańosa (Bradley, 2005, 1977). En realidad, la fisiología del sueńo es tan compleja como la de la vigilia en su regulación neurológica, endocrina, metabólica o cardiorrespiratoria. El estado de sueńo puede sufrir alteraciones por motivos físicos o psíquicos, que ocasionen trastornos potencialmente graves.
En el sueńo se precisa de un ambiente y
una postura adecuados, que son variables en distintas especies:
hay animales que pueden dormir de pie, y otros que pueden hacerlo con
los ojos abiertos.
En contraposición al coma, el estado de
sueńo es reversible en respuesta a estímulos adecuados y genera cambios
electroencefalográficos que lo distinguen del estado de vigilia (Guyton, 2005, 831).
La disminución en la motricidad de la musculatura esquelética y en el
umbral de reactividad a estímulos son otras dos características de este
estado.
El sueńo es periódico y en general
espontáneo, y se acompańa en el hombre de una pérdida de la conciencia
vigil.
Sin embargo, aun cuando el hombre tenga sueńo, puede, voluntariamente,
no dormir. El sueńo tiene distintos grados de profundidad, y se
presentan modificaciones fisiológicas concretas en cada una de las
etapas del mismo (McCarley, 1995, 554.)
Para el estudio de los cambios
funcionales que se dan durante el sueńo se atiende a unas variables que
se denominan indicadores del sueńo:
· El
electroencefalograma (EEG)
· Los
movimientos oculares
· El tono
muscular
La polisomnografía es el registro
de los tres indicadores (Moizeszowicz, 1998, 388)
.
TEORÍAS BÁSICAS SOBRE EL SUEŃO
Se cree que el sueńo está producido por
un proceso inhibidor activo. Una de las primeras teorías del sueńo
proponía que las áreas excitadoras del tronco encefálico, el sistema
reticular activador, se fatigaban a lo largo del día de vigilia y por
tanto se inactivaban: TEORÍA PASIVA DEL SUEŃO (Bremmer, 1935).
Un experimento cambió este punto de vista
por la creencia de que el sueńo probablemente está producido por un
proceso inhibidor activo. Se descubrió que la sección de tronco
encefálico por la región protuberancial media crea un cerebro cuya
corteza nunca duerme (Moruzzi et al, 1949). En
otras palabras, parece existir un centro del sueńo debajo del nivel
protuberancial medio que inhibe otras partes del cerebro.
Según estos indicadores, se distinguen
varias etapas en el sueńo:
El Insomnio Familiar Grave es una
enfermedad de tipo priónico y evolución fatal que fue descrita por
primera vez en los ańos ochenta, cuyo estudio permitió descubrir la
importancia de tal estructura talámica para la instauración del sueńo
lento o profundo (Lugaresi, 1986, 1003).
Es la fase del sueńo más reparadora. Hay movimientos organizados del
dorso; el individuo da vueltas en la cama, cambia de postura. Esta fase
dura aproximadamente un 25% del total del tiempo del sueńo.
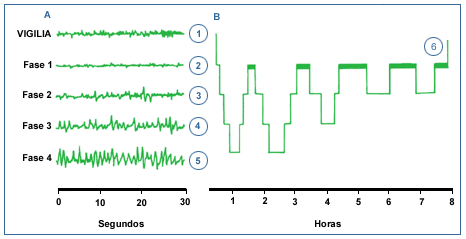
Las etapas I a IV se denominan en su conjunto sueńo NO REM (NREM).
La siguiente etapa es la de sueńo
paradójico, que se caracteriza por una actividad EEG que recuerda
al estado de vigilia (por eso se habla de sueńo paradójico), debida a
una activación cortical por parte de estructuras encefálicas profundas,
como es la formación reticular activadora. Fue descubierto por Kleitman
y Aserinsky, junto con Dement, en los ańos cincuenta del siglo pasado.
Hay una desincronización del EEG, que se asemeja a una situación de
vigilia, de alerta. Se observan movimientos oculares rápidos (también
se habla de sueńo MOR, de movimientos oculares rápidos o sueńo REM, de
rapid eye movements), dependientes de la actividad de estructuras
profundas tales como la formación reticular pontina
(Aserinsky, 1953, 273 ).
Se produce una atonía (desaparición del
tono muscular), de lo que son responsables estructuras como la
formación reticular bulbar, el locus coeruleus, etc. El músculo
diafragma sigue manteniendo el tono, y contrayéndose, permitiendo la
respiración.
La regulación de la vigilia y del sueńo implica al conjunto del SNC, aunque ciertas áreas tienen una importancia crítica.
En el tronco cerebral, diencéfalo y prosencéfalo basal, existen centros cuya influencia es contrapuesta sobre el tálamo y la corteza cerebral; cuando predomina el sistema activador reticular el individuo está alerta, y cuando su influencia decae los sistemas inhibidores inducen el estado de sueńo (Guyton, 2005, 835).
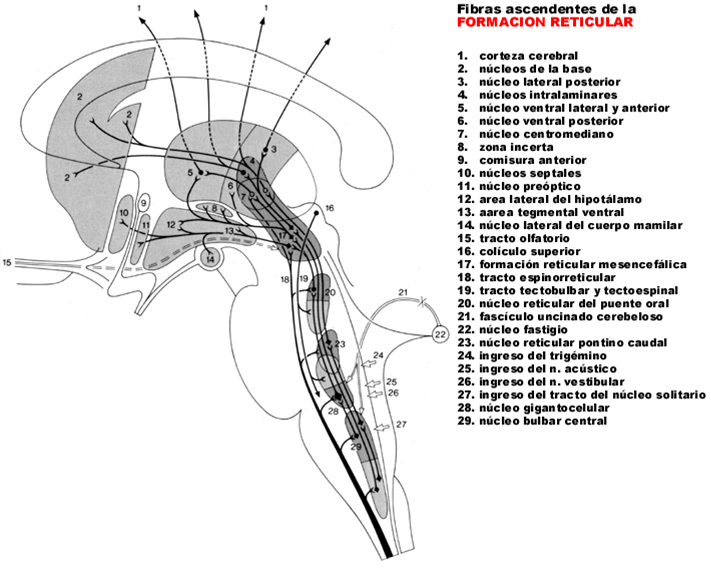
El proceso del ciclo vigila-sueńo está
regulado por una red neuronal compleja en la que intervienen diversas
zonas del sistema nervioso central, a base de activaciones y de
inhibiciones, cuyo resultado es la vigila o el sueńo. Dentro del sueńo,
la fase de sueńo REM es regulada por una complicada red neural en la
que intervienen diversos neurotransmisores.
(Velayos et al., 2007, 8).
Una explicación simplificada no es
posible (Fig. 3).

El mantenimiento de la Vigilia se
debe, sobre todo, a la actividad tónica de las neuronas
catecolaminérgicas y colinérgicas del sistema reticular activador.
También facilitan el estado de vigilia proyecciones histaminérgicas y
peptidérgicas del hipotálamo posterior. La actividad de los sistemas
sensitivos y sensoriales (visual, auditivo) también contribuye al
mantenimiento de la vigilia. A través del área postrema del bulbo,
donde la barrera hematoencefálica es menos activa, algunas sustancias
del torrente sanguíneo como la adrenalina pueden contribuir a la
activación del sistema reticular (Cambier, 2000,
123).
En la génesis del sueńo lento o
NREM intervienen de manera decisiva los núcleos serotoninérgicos
del rafe del tronco cerebral, así como el núcleo del fascículo
solitario, el núcleo reticular talámico, el hipotálamo anterior y
núcleos del área preóptica y el prosencéfalo basal. Las neuronas
serotoninérgicas bloquean la actividad motora y la intensidad de las
aferencias sensoriales. Otros neurotransmisores inhibidores son la
adenosina y el ácido g-aminobutírico (GABA), así como diversos
péptidos. La desactivación progresiva del sistema colinérgico reticular
activador permite la aparición de los ritmos recurrentes
talamocorticales que dan origen a los «husos de sueńo» y al
enlentecimiento del EEG (Zarranz, 2001, 681)
.
La regulación del sueńo REM es aún
más compleja, pues en él se producen al mismo tiempo fenómenos
fisiológicamente antagónicos, como la disminución profunda de la
vigilancia con un estado de activación del EEG o la intensa inhibición
motora con hipotonía generalizada junto con movimientos rápidos
oculares y otras actividades motoras fásicas (distintas en las
diferentes especies) (Ojeda et al., 2004, 249)
.
Todos estos fenómenos están regulados por
diferentes núcleos del tronco cerebral: (Rouviečre,
2005, 255)
Aún se ignora en gran medida cuál es la
finalidad fisiológica última que impone la necesidad que tienen todos
los animales de dormir a pesar de que ello los expone a múltiples
riesgos. El sueńo es una necesidad absoluta y su supresión total
durante suficiente tiempo produce graves trastornos e incluso la muerte
en los animales de experimentación.
Una supresión total de sueńo en el hombre
se tolera sólo si es breve, de 1-2 noches. Si persiste más tiempo
aparecen trastornos progresivamente graves de la atención, la
concentración y la memoria, rritabilidad, alucinaciones e incluso
convulsiones (Pocock et al., 2005, 205).
Una supresión parcial de sueńo puede tolerarse mucho tiempo, a veces a
costa de fatiga crónica.
El registro combinado del EEG, el
electrooculograma, el electromiograma de los músculos submandibulares,
el ECG y la respiración (lo que se conoce como poligrafía del sueńo o
hipnograma) constituye la base objetiva para el estudio del sueńo (Bradley, 2005, 2021) .
Normalmente, en el adulto, el sueńo
nocturno de unas 8 horas se organiza en 4-5 ciclos de unos 90-120 min
durante los cuales se pasa de la vigilia (estadio o fase I) a la
somnolencia (fase II), al sueńo lento (fases III y IV) y finalmente al
sueńo REM (fase V) (López et al., 2005, 668)
.
Su distribución estándar en un adulto
sano es aproximadamente la siguiente:

Las necesidades de sueńo son muy
variables según la edad y las circunstancias individuales (Pocock et al., 2005, 205).
El sueńo es un estado dinámico, en el que
se activan e inhiben distintas zonas del encéfalo, con las
repercusiones funcionales que se han indicado. Obedece a un ritmo
biológico, circadiano (cada 24 horas), relacionado con el ritmo día-
noche, nictemeral, al que se ajusta el proceso
(Pocock, 2005, 206).
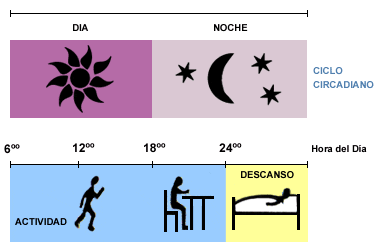
De por sí, el ritmo sueńo-vigilia es cada
25-29 horas, según se ha estudiado experimentalmente en voluntarios
encerrados en una habitación a la que no llegan las influencias
exteriores. Pero la presión del sueńo aumenta en torno a las dos de la
tarde, lo que explica que sea fisiológico sentir sueńo después de
comer. La voluntad puede evitar el dormir después de comer.
Existen unos relojes biológicos en el
sistema nervioso central. Uno de ellos, situado en el hipotálamo
(núcleo supraquiasmático), establece el ritmo sobre los otros
relojes biológicos, situados caudalmente, y hace que el sueńo NREM y el
sueńo REM duren un tiempo fijado (Ojeda, 2004,
273). Intervienen en su regulación no sólo, y de modo
fundamental, los impulsos retinianos, sino también otras influencias,
como es el pH de la sangre o la glucemia. El núcleo supraquiasmático no
es responsable en sí mismo del ritmo vigilia - sueńo, pero sí forma
parte de las redes neurales implicadas en el proceso, redes neurales
sobre las que deben de actuar diversos sistemas para hacer que el
proceso quede anulado por un tiempo, y al fin y al cabo retrasado.
Los ritmos circadianos dependen de la
interacción de los estímulos externos, el más importante de los cuales
es la luz, y de estructuras internas que actúan como marcapasos» de la
entrada en juego de las diferentes funciones.
En el individuo completamente aislado de
influencias exteriores (en una cueva) sin modificaciones de temperatura
ni de luz y sin indicaciones externas de horarios convencionales
(trabajo, comidas), los ciclos de sueńo se van alargando a medida que
entra en juego su «marcapasos» interno de manera autónoma.
En condiciones normales, el indicador
externo más poderoso para sincronizar los ritmos circadianos es la hora
de despertarse y levantarse, que puede fijarse estrictamente. La hora
de irse a la cama también es importante, pero la de dormir no puede
fijarse a voluntad. La estructura cerebral decisiva en la organización
de estos ritmos circadianos, neurológicos, metabólicos y endocrinos, es
el núcleo supraóptico del hipotálamo (Ojeda,
2004, 274)., cuya lesión altera profundamente el
acoplamiento de la vigilancia, el sueńo, el apetito, la temperatura, la
secreción hormonal y otros ritmos.
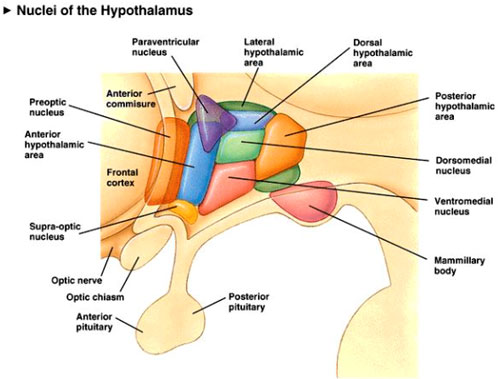
La hormona del crecimiento, el cortisol y
otras hormonas se segregan durante la noche. Así ocurre también con la
melatonina, cuya secreción por la glándula pineal es inhibida por la
luz; al disminuir la luz se incrementa la liberación de melatonina (Cordova, 2003, 717). Ésta favorece la
entrada en el sueńo, quizá permitiendo el descenso de la temperatura
corporal que va ligado a la entrada en sueńo NREM. La exposición
prolongada a la luz, por ejemplo en los viajes transoceánicos en
dirección oeste, inhibe la secreción de melatonina y puede ser la causa
del síndrome del jet lag.
Durante el sueńo hay actividad mental: se
trata de las experiencias oníricas, los ensueńos, que pueden relatarse
en una proporción del 80% al despertar al sujeto después de pasada una
fase de sueńo paradójico.
Por lo tanto, el sueńo no significa falta
de vida ni falta de actividad mental.
Los sueńos de la fase REM son de
tipo cinematográfico y en color. Algunos piensan que el movimiento de
los ojos en esta fase REM va en seguimiento de las imágenes visuales
del ensueńo.
También se sueńa durante la fase de
sueńo profundo, pero estos sueńos son más bien de tipo
abstracto.
El procesamiento de las imágenes de los
ensueńos, según los hallazgos clínicos, se hace de forma preferente en
la corteza occipitotemporal.
En el sueńo REM habría una activación del
sistema reticular activador del tronco del encéfalo y del prosencéfalo
basal, estructuras que también están activas en la vigilia. Estructuras
como los núcleos talámicos relé, que son los que reciben la
estimulación sensorial, quedarían activados, lo que contribuiría a la
aparición de las diversas modalidades psicológicas que aparecen en los
sueńos (Hobson, 1995, 1375).
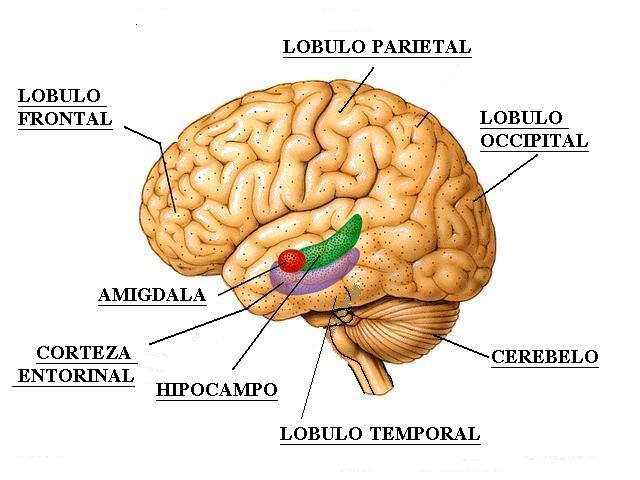
Las estructuras límbicas, tales como la
amígdala y la corteza cingular, también estarían activadas, lo que
explicaría los fenómenos emotivos durante la fase de sueńo REM.
Los ganglios basales y el cerebelo, que
regulan la motricidad, también activados, explicarían los movimientos
ficticios de los sueńos en la fase REM.
Están activas cortezas
asociativas tales como las del lobulillo parietal inferior y de la
corteza occipitotemporal (Orrison, 2000, 150)
, todo lo cual explica las imágenes visuales de esta fase del
sueńo.
La corteza prefrontal, importante
en los procesos mentales, está inhibida, lo cual puede explicar la
falta de lógica en los razonamientos que se experimentan en el sueńo.
Precisamente, según Reinoso (2002), las mismas estructuras que son
necesarias para la consolidación de la memoria, tales como el tálamo
medial, la amígdala, el hipocampo, las estructuras parahipocampales, la
corteza orbitofrontal y cortezas asociativas monomodales están activas
en el sueńo REM.
En el sueńo NREM, en cambio, hay
una inhibición de las zonas que están activadas en el sueńo REM y
activación de las inhibidas, lo que puede explicar que en tal sueńo
NREM haya ensońaciones más de tipo abstracto que en el sueńo REM (Bradley, 2005, 1981). El despertar en
esta fase da lugar a estados confusionales y un rápido retorno al sueńo.
Se puede resumir que el sueńo es un
estado fisiológico activo, en que participan diversas estructuras
encefálicas, formando una red neuronal, en que se dan activaciones e
inhibiciones complejas, con una regulación cíclica, y sobre la que
puede actuar la voluntad, modulando y estableciendo determinadas
actitudes, comportamientos.
Por ejemplo, cada persona tiene
su "liturgia", sus hábitos para dormir: unos necesitan leer un poco
antes de conciliar el sueńo; otros han de tomar un vaso de agua; otros,
precisan de una oscuridad total; otros, de cierto grado de luminosidad,
etc.
Los sueńos son distintos de unas
personas a otras; así por ejemplo, las personas creativas tienen sueńos
particularmente ricos, mientras que las más simples, los tienen más
elementales. Por tanto, la impronta personal también deja su huella en
estos procesos.
Aserinsky E; Kleitman N. Regularly occurring
periods of eye motility, and concomitant phenomena, during sleep.
Science 1953; 118: 273-274.
Belmar J. Estructura, desarrollo
y funciones del sistema nervioso. (en linea) Disponible en:
http://www.puc.cl/sw_educ/neurociencias/
Bradley W. G. 2005. Neurología
Clínica: Diagnóstico y Tratamiento. Madrid: Elsevier
Espańa.
Bremer F. Cerveau "isolé" et
physiologie du sommeil. C.R. Soc. Biol. 1935; 118: 1235-1241.
Cambier J. et al. 2000.
Neurología. Barcelona: Elsevier Espańa.
Cordova A. 2003. Fisiología
dinámica. Barcelona: Elsevier Espańa.
Damiani, D. (2004). Liga de
neurocirugía sistema nervoso. (en linea). Disponible en: http://www.sistemanervoso.com/
Fig. 7. Depresiones (2008). (en linea)
Disponible en:
http://www.depresiones.cl/
Gobierno de Navarra (2008).
Departamento de salud. (en linea) Disponible en:
http://www.cfnavarra.es/salud/anales/default.html
Guyton et al. 2006. Tratado de
fisiología médica. Barcelona: Eselvier Espańa.
Haines. 2003. Principios de
neurociencias. Barcelona: Elsevier Espańa
Hobson JA; Stickgold R. The
conscious state paradigm: a neurocognitive approach to waking, sleeping
and dreaming. En: M. Gazzaniga. The cognitive neurosciences.
Cambridge, Mass., 1995: 1373-1389.
López J. J. et al. 2002. DSM-IV-TR:
Manual diagnóstico y estadístico de los trastornos mentales.
Barcelona: Masson Eselvier.
Lugaresi E; Medori R; Montagna P;
Baruzzi A; Cortelli P; Lugaresi A; et al. Fatal familial insomnia
and dysautonomia with selective degeneration of thalamic nuclei. N
Engl J Med 1986; 315: 997-1003.
McCarley RW. Sleep, dreams and states
of consciousness. En: P. M. Conn. Neuroscience in medicine.
Philadelphia, 1995: 535-554.
Moizeszowicz J. 1998.
Psicofarmacología psicodinámica IV. Estrategias terapéuticas y
psiconeurobiológicas. Barcelona: Elsevier Espańa.
Moruzzi G; Magoun HW. Brain stem
reticular formation and activation of the EEG. 1949. J
Neuropsychiatry Clin Neurosci 1995; 7: 251-267.
Ojeda Sahagún J. L. et al. 2004.
Neuroanatomía humana. Barcelona: Elsevier Espańa.
Orrison W. W. 2000.
Neurorradiología. Madrid: Harcourt.
Pocock G. & Richards C. D. 2005.
Fisiología humana: la base de la medicina. Barcelona: Elsevier
Espańa.
Rodés J. et al. 2004. Medicina
interna. Barcelona: Elsevier Espańa.
Rouvičre H. 2005. Anatomía humana:
descriptiva, topográfica y funcional. Barcelona: Elsevier Espańa.
Vallejo Ruiloba J. 2006. Introducción
a la psicopatología y a la psiquiatría. Barcelona: Elsevier Espańa.
Velayos JL; Reinoso-Suarez F.
Prosencephalic afferents to the mediodorsal thalamic nucleus. J
Comp Neurol 1985; 242: 161-181
Zarranz J. 2001. Compendio de
neurología. Barcelona: Elsevier Espańa.
Nació el 17 de marzo de 1927 en Mecina
Bombarón. A comienzos de los ańos cuarenta se trasladó a Granada para
hacerse maestro y luego cursar la licenciatura de Medicina. Su
inclinación por la morfología comienza cuando gana por oposición el
puesto de Alumno Interno Pensionado de Anatomía y Técnica Anatómica de
la Universidad de Granada.
Su tesis doctoral, sobre el
núcleo rojo, le conducirá a un caminar con huella por el sistema
nervioso. Durante la década de los cincuenta armoniza su formación
morfológica y funcional con estancias en el Instituto Max Planck en
Göttingen y en Giessen y en el Brain Research Institute de Los
Ángeles, al tiempo que madura su carrera universitaria hasta ganar la
cátedra de Anatomía en la universidad de Salamanca en 1957.
Posteriormente ocupa la misma cátedra en las universidades de Granada y
Navarra y, finalmente, desde 1969 a 1992, será catedrático de Anatomía
y Neurobiología y primer director del Departamento de Morfología, de la
Universidad Autónoma de Madrid. En la actualidad es profesor emérito de
Anatomía Humana y Neurociencia de esa universidad.
La estrategia investigadora que
siguió desde los comienzos, los temas de investigación y las
iniciativas docentes le condujeron a ser uno de los impulsores de la
Neurociencia en Espańa desde finales de la década de los cincuenta.
Anima y preside las Reuniones de Neurobiólogos Espańoles y funda, y
también preside, la Sociedad Espańola de Neurociencia. Ha trabajado en
la directiva de la International Brain Organization y varias sociedades
internacionales, y su laboratorio forma parte de la World Federation of
Sleep Research Societies.
Sus trabajos de investigación
sobre el sistema nervioso son esenciales para comprender las relaciones
de las estructuras del tronco del encéfalo con la corteza cerebral, las
conexiones entre distintas áreas corticales y la comprensión de los
mecanismos que gobiernan el sueńo. Su atlas estereotáxico es un
clásico en los laboratorios de neurociencia. Además, con sus
investigaciones ha delimitado morfológica y funcionalmente la región
del tronco del encéfalo donde se origina el sueńo paradójico que es la
fase del sueńo en la que ocurren los sueńos narrativos. Las
investigaciones que ha realizado durante medio siglo son relevantes
para la Neurociencia y, también, para la Psicología, al aportar
conocimientos esenciales del substrato neural del sueńo, la memoria y
las emociones. Su actividad científica se plasma en más de doscientos
trabajos publicados en las revistas más influyentes de Neurociencia y
en revistas generales como Science.
En el curso de su vida académica
ha sido distinguido con múltiples premios y condecoraciones, es
Académico de la Real Academia de Medicina y doctor honoris causa por
las universidades de Granada y Valladolid.