FUNCIONAMIENTO HIPOTALAMICO
(Para Adela, una vez más.Más que nunca.)
RESUMEN
En este documento se intenta explicar el funcionamiento básico del hipotálamo, apoyándose en la anatomía para poder visualizar sus importantes funciones de regulación corporal y cerebral. Para poder describir todo esto se indica todos los núcleos implicados además de los haces nerviosos entre diferentes estructuras cerebrales para poder comprender la complejidad de sus funciones nerviosas además de sus funciones humorales ya que es uno de los principales “órganos” endocrinos y el primer regulador de la secreción hormonal.
PALABRAS CLAVE : Anatomía y Fisiología del hipotálamo
INTRODUCCION
El hipotálamo es uno de los grupos celulares más rostrales de cuantos influyen en las funciones viscerales, y el único con actividad directa sobre el resto de los núcleos visceral es del sistema nervioso central. Además de su papel regulador de las funciones autónomas, también actúa sobre los circuitos nerviosos que modifican el comportamiento. El hipotálamo es el componente diencefálico que interviene en el control central de las funciones viscerales (a través de los sistemas autónomo y endocrino) y del comportamiento afectivo o emocional (a través del sistema límbico). Aunque su cometido fundamental sea el mantenimiento de la homeostasis*, en parte también regula numerosas funciones, como el equilibrio hidroelectrolítico, la ingestión de comida, la temperatura, la presión arterial, tal vez el mecanismo del sueńo-vigilia, los ritmos circadianos así como el metabolismo general del organismo. El hipotálamo es muy pequeńo (unos 4 g) comparado con el resto del encéfalo (unos 1.400 g). Sin embargo, quizá sean los 4 g más importantes de todo el cuerpo. En síntesis, influye sobre nuestras respuestas a los medios interno y externo y es necesario para la vida.
LOCALIZACION ANATOMICA
Límites del hipotálamo
El límite rostral del hipotálamo es la lámina terminal, una delgada membrana que se extiende ventralmente desde la comisura anterior hasta el borde rostral del quiasma óptico. La lámina terminal separa el hipotálamo de los núcleos septales, que quedan por delante. En su parte posterior (dorsal), está limitado por el surco hipotalámico, un surco poco profundo que lo divide del tálamo. Su límite lateral está formado rostralmente por la sustancia innominada y caudalmente por el borde medial del brazo posterior de la cápsula interna. Medialmente, está limitado por la porción inferior del tercer ventrículo. Caudalmente no hay un límite claro; se continúa con la calota mesencefálica y con la sustancia gris periacueductal.
Divisiones del hipotálamo
El hipotálamo se puede dividir en el área preóptica y las zonas lateral, medial y periventricular. El área preóptica es una región de transición que se extiende en sentido rostral hasta continuarse con las estructuras del prosencéfalo basal, por fuera de la lámina terminal. Caudales al área preóptica se localizan otras tres zonas. La delgada zona periventricular es la más medial y se encuentra bajo las células ependimarias que revisten al tercer ventrículo. La zona medial se sitúa lateral a la zona periventricular, y se separa de la zona lateral mediante una línea trazada desde el fórnix poscomisural hasta el fascículo mamilotalámico.
Área preóptica. El área preóptica, aunque funcionalmente forme parte del hipotálamo (y del diencéfalo), embrionariamente deriva del telencéfalo. Está compuesta sobre todo por los núcleos preópticos medial y lateral. El núcleo preóptico medial contiene neuronas que fabrican hormona liberadora de gonadotropinas (GnRH). Esta sustancia se transporta a lo largo del fascículo tuberoinfundibular hasta los capilares del sistema porta hipofisario y, desde allí, hasta el lóbulo anterior de la hipófisis, donde provoca la liberación de las gonadotropinas (hormona luteinizante y hormona foliculoestimulante). Dado que esta liberación es continua en los hombres y cíclica en las mujeres, el núcleo preóptico medial del varón tiende a ser más activo y, por consiguiente, mayor que el de la mujer. Así pues, a menudo se le denomina núcleo sexualmente dimórfico del área preóptica. También influye sobre los comportamientos relacionados con la alimentación, las actividades reproductoras y la locomoción. El núcleo preóptico lateral se localiza inmediatamente rostral a la zona hipotalámica lateral. Aún no se ha determinado totalmente su función; sin embargo, por sus conexiones con el pálido ventral, puede tomar parte en la regulación de la locomoción. Algunos investigadores consideran que los núcleos del área preóptica pertenecen a la región supraóptica de la zona hipotalámica medial. (Haines, 2004, p.480)
Zona lateral. La zona lateral contiene un gran haz de axones, denominado en su conjunto fascículo telencefálico medial. Este haz de fibras difuso atraviesa la zona hipotalámica lateral e interconecta el hipo tálamo con áreas rostrales, como los núcleos septales, y caudales, como la formación reticular del tronco del encéfalo. La zona hipotalámica lateral comprende una gran población difusa de neuronas, llamada habitualmente área hipotalámica lateral, así como unas condensaciones más pequeńas de células localizadas en sus porciones anteriores (Genesser,2006,p.582). Estos últimos grupos son el núcleo hipotalámico lateral y los núcleos tuberales. El núcleo hipotalámico lateral es un conjunto mal definido de células relativamente grandes que se extiende a lo largo de toda la longitud rostrocaudal de la zona hipotalámica lateral. Este núcleo constituye un «centro de la alimentación». Su estimulación en animales de experimentación favorece el comportamiento alimentario; su destrucción hace que se atenúe este comportamiento, y el animal adelgaza. Los núcleos tuberales constan de conglomerados neuronales de escaso tamańo, cada uno formado por pequeńas células pálidas multipolares. Algunas de estas neuronas proyectan hacia el fascículo tuberoinfundibular y, por tanto, pueden transportar hormonas liberadoras hacia el sistema porta hipofisario (Haines, 2004, p.480). Otras envían un estímulo histaminérgico hacia el cerebelo, que puede intervenir en la regulación de la actividad motora.
Zona medial. La zona medial es un sector muy rico en células, compuesto por múltiples núcleos independientes. Se divide en tres regiones: la región supraóptica (quiasmática), la región tuberal y la región mamilar. La región supraóptica se sitúa posterior (dorsal) al quiasma óptico. La región tuberal es la parte más amplia del hipotálamo y corresponde, en general, a la posición del tuber cinereum. La región mamilar, la más posterior de las tres, se encuentra en la zona de los tubérculos mamilares. La región supraóptica está formada por cuatro núcleos: los núcleos supraóptico, paraventricular, supraquiasmático y anterior. Las neuronas de los núcleos supraóptico y paraventricular contienen oxitocina y hormona antidiurética (ADH) (es decir, vasopresina), y transportan estas sustancias hasta la neurohipófisis a través del fascículo supraopticohipofisario, para su liberación al sistema circulatorio. El núcleo supraquiasmático recibe seńales directas desde la retina y tiene capacidad de influir sobre otras estructuras hipotalámicas, como el núcleo preóptico medial. Parece que puede tomar parte en los ritmos circadianos, que son fluctuaciones hormonales secundarias a los ciclos de luz y oscuridad. El núcleo anterior se localiza inmediatamente caudal al área preóptica. La región tuberal contiene tres núcleos: los núcleos ventromedial, dorsomedial y arcuato. El núcleo ventromedial, uno de los más grandes y mejor delimitados del hipotálamo, se considera un «centro de la saciedad». Si se estimula en el laboratorio, el animal de experimentación se despreocupa de los comportamientos alimentarios. A la inversa, su lesión hace que el animal coma demasiado y engorde. El núcleo dorsomedial, localizado inmediatamente posterior (dorsal) al núcleo ventromedial, cumple un papel relacionado con la emoción o, al menos, con el comportamiento afectivo. En los animales de experimentación, su activación despierta conductas desmesuradamente agresivas, que sólo duran el tiempo que se mantenga la estimulación. Este fenómeno, conocido como falsa ira, también se puede provocar mediante la activación de otras zonas hipotalámicas y extrahipotalámicas. El núcleo arcuato es el asiento principal de las neuronas que contienen hormonas liberadoras (Haines, 2004, p.480). Estas sustancias llegan hasta la adenohipófisis a través del fascículo tuberoinfundibular y del sistema porta hipofisario, y allí influyen sobre la liberación de diversas hormonas hipofisarias. La región mamilar consta de cuatro núcleos: los núcleos mamilares medial, intermedio y lateral, y el núcleo hipotalámico posterior. El núcleo mamilar medial es grande y está especialmente bien desarrollado en la especie humana. Representa el punto de terminación principal para los axones del fórnix poscomisural, que se originan sobre todo en el subículo del complejo hipocámpico. También es la fuente de los axones que se dirigen hacia los núcleos anteriores del tálamo formando el fascículo mamilotalámico. Esta última vía constituye una parte importante del sistema límbico. Los núcleos mamilares intermedio y lateral, mucho más pequeńos, quedan laterales al núcleo mamilar medial. El núcleo mamilar lateral recibe estímulos desde la zona medial de la formación reticular mesencefálica a través del pedúnculo mamilar.
Zona periventricular. La zona periventricular, que no ha de confundirse con el núcleo paraventricular es una región muy delgada compuesta por pequeńos somas celulares, que se sitúa por dentro de la zona medial y justo debajo de las células ependimarias del tercer ventrículo. Muchas neuronas de esta zona sintetizan hormonas liberadoras. Sus proyecciones se dirigen hacia el sistema porta hipofisario a través del fascículo tuberoinfundibular, para influir sobre la liberación de diversas hormonas por parte de la adenohipófisis. Por consiguiente muchas de las células de la zona periventricular cumplen una función semejante a la de las neuronas del núcleo arcuato.
CONEXIONES AFERENTES Y EFERENTES
Fibras aferentes al hipotálamo
El hipotálamo está conectado con diversas zonas, como el hipocampo, la amígdala, la calota del tronco del encéfalo, varios núcleos talámicos, los núcleos septales e incluso con áreas neocorticales como la corteza infralímbica y cingular. Con muy pocas excepciones, estas conexiones son recíprocas. A continuación, se ofrecen las entradas y salidas de información más importantes en el hipotálamo.
Fórnix. El fórnix nace a partir de las neuronas del subículo y el hipocampo (dos componentes del complejo hipocámpico) y es la mayor de las vías que llegan al hipotálamo. A medida que se acerca a la comisura anterior, se divide en un pequeńo haz precomisural, derivado fundamentalmente del hipocampo, y un gran haz poscomisural, procedente sobre todo del subículo. El primero se dirige hacia los núcleos septales y preópticos así como hacia la región hipotalámica anterior, mientras que el segundo proyecta básicamente al núcleo mamilar medial, con menos fibras hacia los núcleos talámicos anteriores y el hipotálamo lateral.
Fascículo telencefálico medial. El fascículo telencefalico medial es una estructura mixta difusa, formada fundamentalmente por fibras que siguen un trayecto rostrocaudal a través de la zona hipotalámica lateral. Contiene axones ascendentes y descendentes que interconectan los núcleos septales (incluido el núcleo accumbens septal), el hipotálamo y la calota mesencefálica. Fibras amigdalohipotalámicas. Desde el complejo amigdalino proyectan dos importantes sistemas de fibras aferentes hacia el hipotálamo: la estría terminal, que es la más antigua filogénicamente, y un haz mixto más moderno llamado vía amigdalófuga ventral. La estría terminal se origina en la porción corticomedial de la amígdala y termina en los núcleos septales, el área preóptica y la zona hipotalámica medial. Este haz, que acompańa a la vena terminal en la unión entre el núcleo caudado y el tálamo, sigue la forma arqueada del primero y, por tanto, lleva un trayecto bastante parecido al del fórnix. La vía amigdalófuga ventral nace en la porción basolateral del complejo amigdalino y se dirige en sentido rostromedial bajo el núcleo lenticular y a través del área de la sustancia innominada, hasta entrar en el hipotálamo. Los axones de esta vía finalizan sobre todo en la zona hipotalámica lateral y en núcleos septales y preópticos. Este haz también contiene fibras que viajan desde el hipotálamo hacia la amígdala.
Otras fibras aferentes. El pedúnculo mamilar es un haz difuso de fibras que se origina en las porciones mediales de la formación reticular mesencefálica y termina básicamente en el núcleo mamilar lateral. Algunos de sus axones se incorporan al fascículo telencefálico medial y proyectan hacia los núcleos septales. El núcleo dorsomedial del tálamo da origen a las fibras talamohipotalamicas, que siguen un recorrido en sentido anterior (ventral) hasta entrar en el hipotálamo lateral.
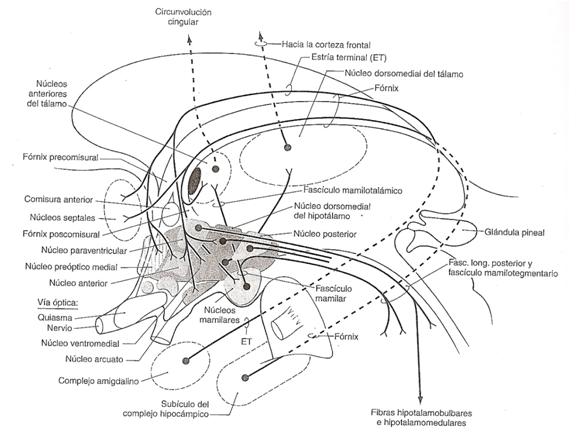 Figura 1. (Haines,
2004, p.485)
Figura 1. (Haines,
2004, p.485)
Fibras eferentes al hipotálamo
Las diversas subdivisiones del hipotálamo poseen proyecciones difusas hacia numerosos lugares a lo largo del sistema nervioso central. Una generalización práctica consiste en tener en cuenta que la mayoría de las estructuras que proyectan hacia el hipotálamo reciben una conexión recíproca desde él. Por ejemplo, al hipotálamo le llega una proyección amigdalohipotalámica y da origen a fibras hipotalamoamigdalinas. Para facilitar su análisis, se dividen las vías de salida según se dirijan hacia estructuras prosencefálicas (ascendentes) y hacia el tronco del encéfalo y la médula espinal (descendentes).
Proyecciones ascendentes. El fascículo mamilar se origina como un haz bien delimitado en el núcleo mamilar medial. Sigue un recorrido posterior (dorsal) durante un corto trayecto y después se bifurca en el fascículo mamilotalámico y el fascículo mamilotegmentario. El primero proyecta hacia los núcleos anteriores del tálamo y es un componente importante del circuito de Papez. El segundo gira en sentido caudal y se distribuye por los núcleos tegmentarios de la formación reticular mesencefálica, como el elemento recíproco del pedúnculo mamilar. Las fibras hipotalamotalámicas surgen fundamentalmente a partir del área preóptica lateral y proyectan hacia el núcleo dorsomedial del tálamo. El hipotálamo también está conectado con los núcleos de la amígdala (fibras hipotalamoamigdalinas) a través de la estría terminal y de la vía amigdalófuga ventral. Estos axones proceden de diversos núcleos hipotalámicos y proyectan sobre todo hacia los núcleos corticomediales del complejo amigdalino.
Proyecciones descendentes. Las principales proyecciones descendentes desde el hipotálamo son cuatro. Se trata de las fibras hipotalamomedulares e hipotalamobulbares, el fascículo longitudinal posterior (dorsal) y el fascículo mamilotegmentario. Las fibras hipotalamomedulares e hipotalamobulbares proceden fundamentalmente del núcleo paraventricular, aunque algunas se originan en los núcleos hipotalámicos dorsomedial y lateral. Estos axones descienden a través de la sustancia gris periacueductal y de la forración reticular adyacente del mesencéfalo y de la porción rostral de la protuberancia, y a continuación se desplazan hacia una posición anterolateral (ventrolateral) en el bulbo raquídeo. Las fibras hipotalamobulbares terminan en el núcleo solitario, el núcleo motor dorsal del vago, el núcleo ambiguo y otros núcleos de la zona ventrolateral del bulbo (Amat, 2001, p.1469). Las fibras hipotalamomedulares recorren la zona anterolateral (ventrolateral) del bulbo y el cordón lateral de la médula espinal para finalizar en las neuronas de la columna celular intermediolateral (células preganglionares motoras viscerales generales). Estos dos grupos de fibras establecen una unión directa esencial entre el hipotálamo y los núcleos autónomos del bulbo raquídeo y de la médula espinal. Pueden quedar interrumpidas por lesión de la zona ventrolateral del bulbo. Aunque no se conocen bien los efectos funcionales de la destrucción de las fibras hipotalamobulbares, la lesión de las fibras hipotalamomedulares ocasiona la pérdida de las respuestas simpáticas en el lado ipsolateral de la cara y de la cabeza (originando un síndrome de Horner), y en el tronco. El fascículo longitudinal posterior (dorsal) se origina en los núcleos de la zona hipotalámica medial, mientras que los axones del fascículo mamilotegmentario nacen en el núcleo mamilar medial. Las fibras de estas vías descienden a través de la sustancia gris periacueductal y acaban fundamentalmente en ella. El fascículo mamilotegmentario, con una posición más anterior (ventral), también acaba en neuronas de los núcleos tegmentarios posterior (dorsal) y anterior (ventral) situados en la sustancia gris periacueductal de la zona caudal del mesencéfalo. Algunas neuronas de esta estructura actúan, al menos en parte, para transmitir la información hacia áreas visceral es del tronco del encéfalo como los núcleos solitario y motor dorsal del vago.
Conexiones intrínsecas
Las vías que interconectan los múltiples núcleos del hipotálamo son numerosas y complejas. El fascículo supraorticohipofisario y el fascículo tuberoinfundibular.
Fascículo supraopticohipofisario. La neurohipófisis libera dos hormonas: oxitocina y hormona antidiurética (ADH). Estas hormonas se sintetizan en las grandes neuronas (magno celulares) de los núcleos supraóptico y paraventricular, llegan a la neurohipófisis a través de sus axones, que forman el fascículo supraopticohipofisario. Las neuronas que contienen oxitocina vierten esta hormona durante el coito, la succión del pezón y los períodos de mayores concentraciones de estrógenos. Esta liberación provoca la contracción del músculo liso uterino y de las células mioepiteliales de la glándula mamaria. Los efectos de la oxitocina sobre el útero resultan decisivos durante el parto y después de él. Tras el nacimiento del bebé, la oxitocina sigue siendo importante. Durante la lactancia, la succión del pezón causa su liberación desde neurohipófisis, y su presencia ocasiona a su vez la contracción de las células mioepiteliales de las glándulas mamarias, con la expulsión de leche. La principal función de la ADH (vasopresina) es contribuir al mantenimiento normal de la osmolaridad sanguínea y la presión arterial. Habitualmente, la ADH eleva la presión arterial, aumentando la volemia. Sin embargo, a valores altos producirá la contracción del músculo liso vascular y también puede originar un ascenso de la presión arterial. A este respecto, su liberación desde la neurohipófisis muchas veces ocurre en situaciones en las que resultaría beneficioso un incremento de la presión arterial. Por ejemplo, la hipotensión que sucede junto con una hipovolemia (descenso del volumen de sangre) representa un estímulo que favorece su liberación. Como resultado, tiene lugar una contracción en las arterias y se eleva la presión arterial. Por tanto, se alivia parcialmente la situación de hipotensión. (Guyton, 2001, p.1028)
Fascículo tuberoinfundibular. La mayoría de los estímulos dirigidos a la hipófisis que llegan a través del fascículo tuberoinfundibular proceden de pequeńas neuronas (parvocelulares) localizadas en el núcleo arcuato y en la zona periventricular. Las neuronas de los núcleos paraventricular, supraquiasmático, tuberal y preóptico medial también contribuyen a formar este fascículo. Los axones transportan varias hormonas liberadoras hacia la eminencia media (la superficie más inferior del área tuberal) y hacia el infundíbulo de la hipófisis. A continuación, estas sustancias se vierten en un plexo primario de capilares fenestrados (sinusoides), desde el que viajan por venas porta hasta otro plexo secundario de capilares fenestrados en la hipófisis. Estos productos hipotalámicos son las diversas hormonas liberadoras (de tirotropina, hormona del crecimiento, corticotropina, gonadotropinas y prolactina), y la hormona inhibidora de la liberación de hormona del crecimiento (somatostatina) .En el lóbulo anterior, las hormonas hipotalámicas regulan el funcionamiento de las células adenohipofisarias productoras de hormonas. Las hormonas de la adenohipófisis son la hormona del crecimiento (que afecta sobre todo al desarrollo del sistema musculo esquelético), las gonadotropinas (que influyen sobre el ovario y el testículo), la corticotropina (que actúa sobre la corteza de la glándula suprarrenal), la tirotropina (dirigida a la glándula tiroides) y la prolactina (con una actividad sobre la producción de leche). Estas hormonas abandonan la adenohipófisis a través de las venas hipofisarias y se distribuyen por la circulación general.
REFLEJOS Y FUNCIONES REGIONALES
Funciones regionales del hipotálamo
Como consecuencia de las interrelaciones existentes entre los sistemas autónomo, endocrino y límbico, y debido a las pequeńas dimensiones del hipotálamo, es difícil asignar una función específica a cada uno de sus componentes individuales. Algunos núcleos hipotalámicos participan en funciones compartidas con otros de su inmediata vecindad. Por consiguiente, estos últimos núcleos pueden intervenir en acciones que no son suyas en exclusiva; en su lugar, pueden actuar de forma concertada para influir sobre funciones atribuibles a regiones hipotalámicas, más que a núcleos específicos. Aunque las acciones de algunos núcleos hipotalámicos se conocen bien, esto no es así en otros casos. Por esta razón y por el hecho de que son muchos los núcleos que toman parte en las funciones regionales, algunos investigadores creen que no se deberían describir «centros» funcionales en el hipotálamo, como un «centro de la alimentación» o un «centro de la saciedad». Además, por las interrelaciones funcionales que existen entre ciertos núcleos dentro de una región hipotalámica dada, merece la pena concebir el hipotálamo según actividades regionales. Las observaciones clínicas coinciden con esta visión. A partir de las observaciones clínicas y los daros experimentales, es oportuno dividir el hipotálamo en dos áreas que comparten funciones semejantes pero opuestas. Estas regiones son las áreas caudolateral y rostromedial. En general, el área caudolateral consta de la zona hipotalámica lateral y la región mamilar, y el área rostromedial de la región supraóptica y gran parte de las regiones tuberales.
Hipotálamo caudolateral. La activación (estimulación) del hipotálamo caudolateral produce manifestaciones que en general se asocian a la ansiedad (Guyton, 2001, p.1028). Se trata de: aumento de la actividad del componente simpático del sistema autónomo, incremento de los comportamientos agresivos, más hambre y elevación de la temperatura corporal (como resultado de la vasoconstricción cutánea y el temblor).
Hipotálamo rostromedial. La activación (estimulación del hipo tálamo rostromedial produce manifestación conductuales que en general se asocian a la obtención de una satisfacción. Consisten en: aumento de la actividad del componente parasimpático del sistema autónomo, incremento del comportamiento pasivo, descenso de la temperatura corporal (debido a la vasodilatación cutánea y la sudoración). La sudoración es un fenómeno singular. Aun cuando las glándulas sudoríparas están inervadas por fibras nerviosas simpáticas, la sudoración es compatible con la función parasimpática en cuanto nos sirve para mantener una temperatura baja. El hecho de que las terminales simpáticas que inervan la mayoría de las glándulas sudoríparas sean colinérgicas, igual que las terminales parasimpáticas que inervan las vísceras, y el que la sudoración se pueda provocar desde el hipotálamo rostromedial.
Reflejos hipotalámicos
Todas las funciones vitales del hipotálamo, como el mantenimiento de la presión arterial, la temperatura corporal y el balance hídrico, están reguladas por reflejos y, no se encuentran sometidas a un control consciente. El medio interno está controlado parcialmente a través de los reflejos hipotalámicos, que se encuentran mediados específicamente por los sistemas autónomo y endocrino.
Reflejo barorreceptor*. El reflejo barorreceptor regula la presión arterial como respuesta a las seńales de los barorreceptores del cayado aórtico y del seno carotideo. Estos receptores se llaman extrínsecos debido a que están fuera del sistema nervioso central. Detectan las variaciones en la presión arterial y transmiten información hacia las neuronas del núcleo solitario en el bulbo raquídeo. Estas neuronas proyectan y activan a las del núcleo motor dorsal del vago, que a su vez conectan con los ganglios terminales en el corazón e influyen sobre frecuencia cardíaca. Un nivel de presión arterial por encima de lo normal activa el núcleo solitario, generando descenso de la frecuencia cardíaca y de la fuerza de contracción del corazón y, por consiguiente, una reducción de la presión arterial; un valor por debajo de lo normal tiene el efecto opuesto. El hipotálamo es capaz de influir sobre el reflejo barorreceptor a través de una vía un poco más compleja. El núcleo solitario contiene células que transmiten información barorreceptora hacia los núcleos hipotalámicos paraventricular, dorsomedial y lateral. A su vez, las neuronas de estas áreas hipotalámicas proyectan hacia el núcleo motor dorsal del vago en el bulbo raquídeo. Por este camino, el hipotálamo puede modular de forma potente la capacidad de respuesta al reflejo barorreceptor.
Reflejo de regulación de la temperatura. El reflejo que mantiene constante la temperatura corporal depende de Las seńales procedentes de neuronas termosensibles especializadas del hipo tálamo (llamadas receptores intrínsecos porque están dentro del sistema nervioso central). Cuando la temperatura de la sangre que llega al hipotálamo se eleva por encima de lo normal, estas neuronas estimulan las regiones del hipotálamo rostral responsables de activar los mecanismos fisiológicos para la disipación del calor: sudoración y vasodilatación cutánea. Estos efectos se encuentran mediados por vías autónomas. A la inversa, si la temperatura sanguínea está por debajo de lo normal, los sensores térmicos estimulan regiones del hipotálamo caudal que activan mecanismos para la conservación del calor (vasoconstricción cutánea mediada por vías autónomas) y para su producción (temblor mediado por las vías reticuloespinales).
Reflejo del equilibrio hídrico. El volumen y la osmolaridad de la sangre se mantienen constantes a través de mecanismos reflejos. A diferencia de los ejemplos anteriores, que son totalmente nerviosos, el reflejo del equilibrio hídrico es neurohumoral; su brazo eferente consta de una seńal hormonal transportada por la ADH. La osmolaridad de la sangre se controla mediante unas neuronas especializadas osmosensibles, localizadas en el hipotálamo anterior cerca de los núcleos preóptico y paraventricular. Las respuestas de estos receptores influyen sobre la liberación de ADH desde las neuronas productoras de esta hormona en los núcleos supraóptico y paraventricular. Cuando la osmolaridad sanguínea es demasiado alta, se libera más ADH y se eleva la reabsorción de agua en los túbulos colectores del rińón. Si la osmolaridad es demasiado baja, se inhibe la liberación de ADH y se reduce la reabsorción renal de agua. Así pues, no se reabsorbe agua hacia la sangre, sino que permanece en la orina. (Guyton, 2001, p.375-376)
PATOLOGIAS CON MAYOR INCIDENCIA
Lesiones del núcleo supraóptico, paraventricular, del fascículo supraopticohipofisario:
Producen un síndrome conocido como diabetes insípida, que se caracteriza por poliuria (aumento de la micción) y polidipsia (aumento del consumo de agua). Este proceso se debe a una escasez de la ADH circulante. Es interesante saber que el etanol genera un descenso de su liberación desde la neurohipófisis. Ésta es la razón por la que el consumo de bebidas alcohólicas tiende a causar una micción abundante, y la deshidratación y sed consiguientes.
Lesión del hipotálamo caudolateral:
Produce manifestaciones típicas opuestas a las ocasionadas por su estimulación. Por ejemplo, el dańo de esta área se traduce en la inhibición de las actividades simpáticas y la reducción de la temperatura corporal.
Lesiones del hipo tálamo rostromedial:
Suelen manifestar comportamientos opuestos a los descritos con su estimulación. Por ejemplo, un dańo en esta área causa la inhibición de las actividades parasimpáticas y el aumento de la temperatura corporal.
Síndrome de Korsakoff
El conocimiento de las funciones de los núcleos mamilares procede de las observaciones experimentales y clínicas. Por ejemplo, las lesiones de los tubérculos mamilares tienden a impedir la conservación de la memoria recién adquirida, de modo que la memoria inmediata o memoria a corto plazo no se transforma en memoria a largo plazo. Un paciente con una lesión mamilar no tiene problemas para recordar los hechos sucedidos meses o ańos antes de su producción. Sin embargo, la memoria de los acontecimientos que ocurrieron después de la misma se encuentra restringida a un tiempo breve (un plazo de minutos), y no se crea una memoria a largo plazo. Como resultado de esta amnesia anterógrada, los pacientes afectados suelen tener graves dificultades para aprender nuevas tareas y para transformar sus experiencias en una memoria a largo plazo. Estas deficiencias específicas de la memoria son características del un proceso ocasionado por la carencia de tiamina* y que se asocia clásicamente al alcoholismo crónico. Los defectos de memoria en este síndrome se deben a la degeneración progresiva de los tubérculos mamilares y de estructuras encefálicas funcionalmente relacionadas con ellos, como el complejo hipocámpico y el núcleo mediodorsal del tálamo. Los pacientes con síndrome de Korsakoff pueden tener problemas para comprender información por escrita y para sostener conversaciones coherentes, debido a que tienden a olvidar lo que acaban de leer o de decir. Un rasgo interesante de este síndrome es la tendencia a la fabulación, es decir, a ensartar los fragmentos memorizados con distintos hechos para crear por síntesis un «acontecimiento» que jamás ocurrió.
Tumores hipofisarios
La hipófisis está vinculada anatómica y fisiológicamente al hipotálamo. Así, es la vía para la manifestación de muchas funciones hipotalámicas. Las hormonas convencionales y las hormonas liberadoras ya fabricadas en el hipotálamo viajan a través de los fascículos supraopticohipofisario y tuberoinfundibular, y se vierten al sistema porta hipofisario. Las pequeńas lesiones hipotalámicas pueden bloquear la elaboración y transporte de estas sustancias, afectando así de forma negativa a las funciones hipofisarias. A semejanza de esto, los tumores (adenomas) que aparecen en la hipófisis pueden invadir con facilidad el hipotálamo vecino, y poner así en peligro sus funciones. Aunque los defectos visuales no sean un tema específico de este tema habría que seńalar su frecuente aparición en pacientes con tumores hipofisarios. Estos defectos pueden poner de manifiesto una lesión del nervio óptico próxima al quiasma, del propio quiasma (fibras directas o fibras que se decusan) o de la cintilla óptica situada detrás de él. Los tumores que aparecen en la hipófisis representan el 12% de los tumores cerebrales primarios. En los estudios necrósicos, la presencia total de adenomas hipofisarios descubiertos casualmente (no diagnosticados) ha oscilado entre el 22,5% y el 3,2%, en función del grosor seccionado de la hipófisis. Los tumores hipofisarios que se diagnostican con mayor frecuencia entre los jóvenes no suelen ser malignos. Los adenomas casuales se detectan con más frecuencia en la quinta, sexta y séptima décadas de la vida. Los tumores hipofisarios se pueden clasificar según sus características secretoras, su tamańo, o su poder invasivo. Los tumores secretores producen en exceso una o más hormonas hipofisarias; la prolactina es la más frecuente entre ellas. Una segunda clasificación se efectúa por su tamańo; los microadenomas son tumores cuyo eje mayor no llega a 1 cm, mientras que cuando superan estas dimensiones transversales se denominan macroadenomas. Una tercera clasificación de estos tumores se fija en su poder invasivo; los tumores invasivos pueden causar una erosión y extenderse hacia la duramadre e incluso hasta el esfenoides.
no producen hormonas y, por consiguiente, muchas veces quedan sin diagnosticar mientras no alcancen un tamańo considerable y ejerzan una presión sobre las estructuras próximas (p. ej., el quiasma óptico y el hipotálamo). Los pacientes con estos grandes tumores hipofisarios tienen síntomas, que suelen consistir en trastornos visuales (en el 60 a 70% de los enfermos) y cefaleas.
En el marco de la clínica, los tumores secretores también reciben habitualmente la denominación de tumores hormonamente activos o tumores hipersecretores. Las manifestaciones clínicas de un tumor secretor consisten en los efectos de la actividad biológica de la hormona hipofisaria específica fabricada en exceso. En los casos de hiperproducción de hormona del crecimiento,los pacientes sufren un aumento descontrolado de su estatura, que se conoce como gigantismo, si el proceso sucede antes del cierre de las placas epifisarias de los huesos largos. Sus músculos pueden ser grandes, pero en realidad son bastante débiles porque contienen una mayor cantidad de tejido conjuntiva que de fibras musculares. Por otra parte, la hiperproducción de hormona del crecimiento tras el cierre de las placas epifisarias genera una enfermedad denominada acromegalia, aludiendo al incremento de los dedos del paciente. Los acromegálicos tienen unos cambios faciales típicos, con alargamiento de la cara, trastornos de la oclusión mandibular y de los diastemas (espacios) en la dentadura inferior. Otras alteraciones son los relieves de los senos frontales y mastoideos*, una nariz bulbosa, labios engrosados y manos y pies muy grandes. Asociadas a estas manifestaciones externas está el excesivo engrosamiento del hueso, la insuficiencia cardíaca secundaria al crecimiento del corazón. Una forma mucho menos frecuente de tumor hipofisario puede derivar de la hiperproducción de tirotropina,con el resultado de un hipotiroidismo o un hipertiroidismo. A menudo, los pacientes sufren alteraciones de la función cardiovascular, así como temblor. Según va evolucionando el tumor, pueden identificarse síntomas como las cefaleas, los trastornos visuales y la disfunción de los pares craneales próximos a la silla turca (es decir, el III, IV Y VI). La alteración de los pares craneales suele afectar en primer lugar al nervio motor ocular externo, y a continuación a los nervios motor ocular común y troclear, a medida que crece el tumor en sentido lateral. Una hiperproducción de corticotropina lleva a la aparición de un hipercorticalismo conocido como enfermedad de Cushing. En este caso, el exceso de hormona adrenocorticotropa procedente de la hipófisis provoca una hipersecreción de cortisol en las suprarrenales, con los rasgos clínicos clásicos de la enfermedad de Cushing. En general, los pacientes afectados tienen una obesidad central, del tronco, con cara de luna llena, hirsutismo facial y una joroba cervical dorsal, habitualmente llamada cuello de búfalo, que deriva del crecimiento de la almohadilla adiposa en esta región. La obesidad centrípeta tiene como dato característico las estrías violáceas (marcas de estiramiento). Estas estrías son claramente diferentes de las que se observan en el embarazo o entre las personas demasiado obesas. En la enfermedad de Cushing son de color morado o violeta, mientras que en otras situaciones suelen ser blancas. Estos pacientes también sufren una hiperpigmentación, hematomas cutáneos, hipertensión, osteopenia e inestabilidad emocional (Guyton, 2001, p.1105) .Los secretores de prolactina hiperproducción de prolactina en las mujeres produce un síndrome de galactorrea (producción de leche) Y amenorrea (ausencia de menstruaciones). La hiperprolactinemia se observa de forma fisiológica como un componente del embarazo normal. Sin embargo, hay otras causas que requieren su diferenciación de un tumor o de una gestación (p. ej., el hipo tiroidismo o el consumo de drogas). En los hombres, puede manifestarse por un descenso de la libido, impotencia o esterilidad. Este tipo de tumor hipersecretor es una causa importante de esterilidad. Tumores gonadótropos. Estos tumores están formados por células que producen una cantidad excesiva de hormona luteinizante (LH) o foliculoestimulante (FSH). La hipersecreción de FSH no causa síntomas conocidos en el hombre o en la mujer posmenopáusica. Se ha seńalado que la de LH ocasiona cambios puberales prematuros (pubertad precoz) en el hombre, y tal vez interrupción de los ciclos ováricos en la mujer. Sin embargo, en la mayoría de los casos los adenomas gonadótropos llaman la atención' en la clínica debido a los efectos secundarios a la masa tumoral, con perturbaciones visuales, cefaleas y a veces diplopía (visión doble) por la compresión del nervio óptico secundaria a la extensión lateral del tumor.
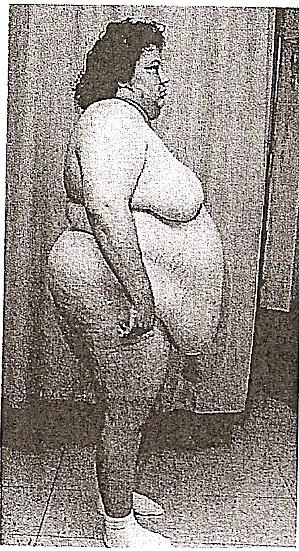 Figura 2. Mujer con la enfermedad
de Cushing debida a la hiperproducción de corticotropina.
(Haines, 2004, p.490)
Figura 2. Mujer con la enfermedad
de Cushing debida a la hiperproducción de corticotropina.
(Haines, 2004, p.490)
BIBLIOGRAFIA
GLOSARIO DE TERMINOS
Baroreceptor
Los barorreceptores son receptores de presión situados en el seno carotideo y en el cayado aórtico. Cuando aumenta la presión arterial aumenta el número de impulsos que envían por el nervio de Hering y por el vago hasta el núcleo del tracto solitario en el bulbo raquídeo.
Tiamina
La tiamina (o Vitamina B1) es una vitamina hidrosoluble que, en forma de pirofosfato, interviene como coenzima en diversas reacciones. Está distribuida ampliamente, tanto en alimentos vegetales como animales, de modo que en principio es suficientemente abundante en todos los alimentos sin procesar. Sin embargo, su localización en los cereales, en la cascarilla en el arroz o en el salvado en el trigo, hace que se pierda un porcentaje importante en su preparación industrial (en el caso del arroz, toda). Además es una vitamina muy lábil.
Senos frontales y mastoideos
Cavidades existentes en el espesor de un hueso o formada por la reunión de varios huesos, en este caos de frontal e independientemente de las apófisis mastoides.
Homeostasis
Proceso por el cual un organismo mantiene las condiciones internas constantes necesarias para la vida.
BIOGRAFIA
Arthur Guyton
(1919-2003). Fisiólogo especializado en patologías cardiovasculares. Sus últimos ańos de vida trabajo en el Hospital Presbiteriano de Nueva york. Su obra más representativa de sus experimentos es Tratado de fisiología médica donde analiza todo el funcionamiento del organismo humano.
Finn Geneser
(1967) Histólogo de la Universidad de Aarhus. Su obra con mayor repercusión es la citada en el trabajo: Histología: sobre bases biomoleculares. Realizada su primera edición en 1981.
INDICE ANALITICO
ADH: 3, 8, 11, 12.
FSH: 15.
Fornix: 2, 4, 5.
Núcleos
Osmolaridad: 8, 11
Oxitocina: 3, 8.